- Dromaeosauridae
-
 Dromaeosauridae
Dromaeosauridae
Deinonychus antirrhopus (le grand)
et Buitreraptor gonzalezorum (le petit)
Field Museum of Natural History, Chicago.Classification Règne Animalia Classe Sauropsida Ordre Saurischia Sous-ordre Coelurosauria Famille Dromaeosauridae
Colbert et Russell, 1969Genres de rang inférieur - Achillobator
- Adasaurus
- Atrociraptor
- Bambiraptor
- Balaur
- Cryptovolans
- Deinonychus
- Dromaeosauroides
- Dromaeosaurus
- Graciliraptor
- Linheraptor
- Microraptor
- Pyroraptor
- Saurornitholestes
- Sinornithosaurus
- Utahraptor
- Variraptor
- Velociraptor
Les Dromaeosauridae (ou dromaeosauridés) sont une famille de dinosaures théropodes ressemblant à des oiseaux. Ils étaient des carnivores à plumes de taille petite à moyenne qui ont abondé au cours du Crétacé. Le nom Dromaeosauridae signifie « lézard coureur », du grec dromeus (δρομευς) qui signifie « courir » et sauros (σαυρος) qui signifie « lézard ».
On a trouvé des fossiles de dromaeosauridés en Amérique du Nord, Europe, Afrique, Japon, Chine, Mongolie, Madagascar, Argentine et Antarctique[1]. Ils sont apparus au milieu du Jurassique (à la fin du Bathonien, il y a environ 164 millions d'années) et ont survécu jusqu'à la fin du Crétacé (Maastrichtien, il y a 65,5 Ma), soit pendant environ 100 millions d'années, jusqu'à l'extinction Crétacé-Tertiaire. La présence de dromaeosauridés dès le Jurassique moyen a été confirmée par la découverte de dents fossiles isolées, mais aucun corps fossilisé de dromaeosauridé de cette époque n'a été retrouvé[2].
Sommaire
Description
 Velociraptor mongoliensis restauré avec ses grandes plumes aux ailes comme on peut le déduire aux points de fixation des pennes
Velociraptor mongoliensis restauré avec ses grandes plumes aux ailes comme on peut le déduire aux points de fixation des pennes
Les caractéristiques du corps des dromaeosauridés ont contribué à ranimer les théories qui voulaient qu'au moins certains dinosaures aient été actifs, rapides et étroitement liés aux oiseaux. L'illustration faite par Robert Bakker pour la monographie de John Ostrom datant de 1969[3], montrant le dromaeosauridé Deinonychus lancé dans une course rapide, est parmi les reconstructions les plus influentes de l'histoire de la paléontologie[4]. Les dromaeosauridés avaient une tête relativement grosse, des dents crénelées, un museau étroit et des yeux orientés vers l'avant ce qui indique un certain degré de vision binoculaire[5]. Les dromæosauridés, comme la plupart des autres théropodes, avaient un long cou en S et un tronc relativement court et haut. Comme les autres maniraptoriens, ils avaient de longs bras qui pouvaient être repliés contre le corps chez certaines espèces et des « mains » relativement importantes avec trois longs doigts (le doigt du milieu étant le plus long et le premier doigt le plus court) se terminant par de grandes griffes[6]. La structure de la hanche des dromaeosauridés avait aussi ce caractéristique évasement du pubis faisant saillie sous la base de la queue. Les pieds des dromaeosauridés portaient une grande griffe courbe au niveau du deuxième orteil. Leur queue était effilée, avec de longues vertèbres étroites, qui perdaient leurs processus transverses et arrière à partir de la 14e vertèbre caudale[6].
On sait maintenant qu'au moins certains, et probablement tous, les dromæosauridés étaient couverts de plumes, y compris de grandes plumes à pinnules au niveau des ailes et de la queue. Cette hypothèse, d'abord émise au milieu et à la fin des années 1980 a été confirmée par des découvertes fossiles en 1999, représentant un changement important dans la façon dont les dromæosauridés ont toujours été représentés dans l'art et le cinéma (voir le paragraphe « plumes » ci-dessous).
Pieds
 Os du pied reconstitué d'un Deinonychus. Le deuxième orteil porte une grande griffe rétractée.
Os du pied reconstitué d'un Deinonychus. Le deuxième orteil porte une grande griffe rétractée.Comme les autres théropodes, les dromæosauridés étaient des bipèdes. Cependant, alors que ces premiers marchaient avec trois doigts en contact avec le sol, les empreintes de pas fossilisées de dromæosauridés confirment que la plupart d'entre eux avaient le deuxième orteil tenu en hyperextension et ne touchant pas terre, le poids de l'animal portant seulement sur les troisième et quatrième rayons. C'est ce qu'on appelle une « didactylie fonctionnelle »[7]. Le second orteil portait une grande griffe en forme de faucille (qu'il tenait écartée du sol lors de la marche) et qui aurait pu servir à capturer des proies et grimper dans les arbres (voir le paragraphe « Rôle de la griffe » ci-dessous). Une espèce de dromaeosaurien, Balaurs bondoc, possédait également un premier orteil fortement modifié un peu comme le second. Ces deux orteils de chaque pied étaient également tenus rétractés et possédaient de grandes griffes en forme de faucille[8].
Queue
Les dromæosauridés avaient une longue queue. La plupart des vertèbres caudales possédaient des extensions osseuses en forme de tige et même des tendons ossifiés chez certaines espèces. Dans son étude sur Deinonychus, Ostrom a proposé que ces traits caractéristiques servaient a raidir la queue de sorte qu'il ne pouvait la plier qu'à la base, et la queue entière se comportait comme un levier rigide[3]. Toutefois, un spécimen bien conservé de Velociraptor mongoliensis (IGM 100/986) avait une queue articulée en forme de S horizontal. Ceci suggère que, dans la vie, la queue pouvait se plier de gauche à droite avec une flexibilité substantielle[10] On a proposé que la queue servait de stabilisateur et/ou de contrepoids quand il courait ou volait[9]. Chez Microraptor, on trouvait un long éventail de plumes à l'extrémité de la queue. Il devait être utilisé comme stabilisateur aérodynamique et organe de direction dans les vols battus ou planés (voir le paragraphe « Vol battu et plané » ci-dessous)[10].
Taille
 Comparaison des tailles de différents dromaeosauridés.
Comparaison des tailles de différents dromaeosauridés.Les dromæosauridés étaient des dinosaures de taille petite à moyenne, allant d'environ 0,7 mètre de longueur dans le cas de Mahakala à plus de 6 m pour Utahraptor et Achillobator[11],[12]. Certains seraient même plus grands ; des spécimens d’Utahraptor non encore décrits figurant dans les collections de l'Université Brigham Young pourraient avoir atteint jusqu'à 11 m de long, mais il faut attendre des études plus détaillées pour en être sûr[13]. L'augmentation de taille semble s'être déroulée au moins en deux fois parmi les dromæosauridés; une première fois chez les dromaeosaurinés Utahraptor et Achillobator, puis une deuxième fois chez l'unenlagiiné Austroraptor qui mesurait 5 m de long. Une troisième augmentation de taille est possible pour des dromaeosaures géants représentés seulement par des dents trouvées sur l'île de Wight, en Angleterre. Les dents appartiennent à un animal de la taille de l’Utahraptor mais qui semble en fait être un vélociraptoriné, à en juger par la forme des dents[14].
Mahakala est à la fois le plus primitif et le plus petit des dromaeosauridés jamais décrit. Ce fait, conjugué à la petite taille des autres dromæosauridés primitifs tels que Microraptor et le troodontidé Anchiornis, indique que l'ancêtre commun des dromæosauridés, troodontidés et des oiseaux - qui est appelé le paravien ancestral - peut-être été très petit, mesurant environ 65 cm de longueur pour un poids de 600 à 700 grammes[15].
Plumes
Article détaillé : Dinosaures à plumes. Moulage de l'holotype de Sinornithosaurus millenii le premier dromaeosauridé découvert avec des plumes
Moulage de l'holotype de Sinornithosaurus millenii le premier dromaeosauridé découvert avec des plumesOn dispose d'un grand nombre de preuves que les dromæosauridés étaient couverts de plumes. Certains fossiles ont gardé leurs longues plumes pennées sur les membres antérieurs (rémiges) et la queue (rectrices), ainsi que de plumes plus courtes, une sorte de duvet, qui recouvraient leur corps[16],[17]. D'autres fossiles, qui n'ont pas conservé traces de plumes, conservent encore sur les os des avant-bras des protubérances où devaient venir s'attacher de longues plumes[18]. Dans l'ensemble, ce modèle de plumage ressemble beaucoup à celui d’Archaeopteryx[16].
Le premier dromaeosaure connu avec certitude pour avoir eu de plumes a été Sinornithosaurus, découvert en Chine par Xu et al. en 1999[17]. De nombreux fossiles d'autres dromaeosauridés ont été trouvés depuis avec des plumes couvrant leur corps, certains avec les plumes des ailes entièrement développées. Microraptor montre même à l'évidence que ses pattes de derrière formaient une seconde paire d'ailes[16]. Bien que les impressions directes de plumes ne soient possibles que dans les sédiments à grains fins, des fossiles trouvés dans des roches à gros grains présentent des signes d'existence de plumes par la présence de points d'insertion de plumes, des points qu'on retrouve chez certains oiseaux. On a trouvé de telles situations chez les dromæosauridés Velociraptor et Rahonavis ce qui prouve que ces deux genres avaient des plumes même si on n'en a pas de preuves directes. Dans ce contexte, il est fort probable que même les plus terricoles des dromæosauridés portaient des plumes, puisque même les oiseaux actuels incapables de voler conservent la plus grande partie de leur plumage et des dromæosauridés relativement importants, comme Velociraptor, sont connus pour avoir conservé leurs plumes pennées[18],[19]. Bien que certains scientifiques aient suggéré que des dromæosauridés avaient perdu une grande partie ou la totalité de leur couverture isolante, la découverte de plumes sur des échantillons de Velociraptor a été considérée comme une preuve que tous les membres de la famille avaient conservé leurs plumes[18],[20].
Paléobiologie
Rôle de la griffe
 Modèle des os du pied d'un dromaeosauridé
Modèle des os du pied d'un dromaeosauridéIl y a actuellement désaccord sur le rôle de la griffe en faucille du deuxième orteil. Lorsque John Ostrom a décrit Deinonychus en 1969, il voyait le rôle de la griffe comme celui d'une lame, d'une arme tranchante, un peu comme les canines de certains tigres à dents de sabre, utilisée en donnant de puissants coups de pied pour couper une proie. Adams (1987) a suggéré que la griffe était utilisée pour éventrer les grands dinosaures cératopsiens[21] . Cette interprétation du rôle de la griffe comme arme à tuer a été appliquée à tous les dromæosauridés. Toutefois, Manning a fait valoir que la griffe devait plutôt avoir un rôle de crochet, la gaine de kératine qui l'entourait ayant une section elliptique, au lieu de la forme de larme inversée précédemment supposée[22]. Dans l'interprétation de Manning, la griffe aurait utilisée comme aide pour grimper sur les grosses proies et aussi comme poignard.
Ostrom a comparé Deinonychus à l'autruche et au casoar. Il a noté que ces espèces d'oiseaux pouvaient infliger de graves blessures grâce à la grande griffe sur leur deuxième orteil[3]. Le casoar a des griffes pouvant atteindre jusqu'à 125 millimètres de longueur[23]. Ostrom cite Gilliard (1958) en disant qu'elles peuvent couper un bras ou éventrer un homme[24]. Kofron (en 1999 et 2003) a étudié 241 documentée sur des attaques de casoar et a constaté qu'un homme et deux chiens avaient été tués, mais qu'il n'existait aucune preuve que des casoars puissent éventrer ou démembrer d'autres animaux[25],[26]. Les casoars utilisent leurs griffes pour se défendre, pour attaquer des animaux menaçants et dans les rivalités de parade en période de reproduction[23]. Le cariama a aussi une griffe au deuxième orteil qu'il l'utilise pour déchirer ses proies en petits morceaux avant de les avaler[27].
Phillip Manning (2009) a tenté de tester le fonctionnement de la griffe des pieds et celles de forme similaire des membres antérieurs. Il a analysé la façon dont les contraintes étaient réparties le long des griffes et des membres, en utilisant un scanner pour créer une carte en trois dimensions d'une griffe de patte antérieure de Velociraptor. A titre de comparaison, il a analysé la conception d'une griffe d'un oiseau prédateur actuel, le Hibou grand-duc. Il a constaté que, selon la façon dont les pressions étaient réparties le long des griffes, elles étaient d'excellents moyens d'escalade. Les scientifiques ont découvert que l'extrémité pointue de la griffe était aussi un instrument de perforation et de préhension, tandis que la courbure et la large base de la griffe aidaient à transférer les contraintes de façon uniforme[28].
L'équipe de Manning a également comparé la courbure de la griffe du pied des dromaeosauridés à la courbure des griffes des oiseaux et des mammifères actuels. Des études antérieures avaient montré que l'importance de la courbure d'une griffe était en corrélation avec le mode de vie de l'animal : les animaux à griffes fortement courbées avaient tendance à être des grimpeurs, ceux à griffes droites un mode de vie terrestre. Les griffes de Deinonychus ont une courbure de 160 degrés, dans la zone des animaux grimpeurs. Les griffes des membres antérieurs qu'ils ont également étudié se trouvent également dans la gamme des animaux grimpeurs[28].
Le paléontologue Peter Mackovicky a commenté l'étude de l'équipe de Manning en disant que s'il admettait que les petits dromæosauridés primitifs (comme Microraptor) aient pu avoir grimpé aux arbres,il ne s'expliquait pas pourquoi, plus tard, les dromæosauridés gigantesques comme Achillobator avaient conservé leurs griffes très incurvées alors qu'ils étaient beaucoup trop lourdes pour grimper aux arbres. Mackovicky pensait tout de même que les dromæosauridés géants pouvaient avoir gardé leur griffe exclusivement pour s'accrocher à leurs proies[29].
En 2009, Phil Senter a publié une étude sur les orteils de dromaeosauridés et montré que leur amplitude de mouvement était compatible avec une excavation de nids coriaces d'insectes. Senter a suggéré que les petits dromæosauridés comme Rahonavis et Buitreraptor étaient assez petits pour être partiellement insectivores, tandis que les grands tels que Deinonychus et Neuquenraptor auraient pu utiliser cette capacité pour capturer des vertébrés résidant dans des nids d'insectes. Toutefois, Senter n'a pas vérifié si la forte courbure des griffes de dromaeosauridés permettait de telles activités[30].
Comportement social
 Empreintes de pas du deinonychosaure Paravipus didactyloides, considérées comme dues à deux individus marchant dans la même direction.
Empreintes de pas du deinonychosaure Paravipus didactyloides, considérées comme dues à deux individus marchant dans la même direction.On a découvert de petits groupes de Deinonychus fossilisés près de restes de Tenontosaurus, un grand dinosaure ornithischien herbivore. On a interprété cela comme une preuve que les dromaeosaures chassaient en meutes comme certains mammifères actuels[31]. Cependant, tous les paléontologues ne considèrent pas cela comme une preuve concluante et une étude subséquente publiée en 2007 par Roach et Brinkman suggère que Deinonychus n'avait peut-être pas de comportement social réellement organisé. Les diapsides actuels, comme les oiseaux et les crocodiles (les plus proches parents des dromaeosaures), ont un minimum de chasse coopérative ; au contraire, ce sont généralement soit des chasseurs solitaires, soit attirés par des carcasses d'animaux précédemment tués et il y a alors souvent des conflits entre individus de la même espèce. Par exemple, lorsque des dragons de Komodo mangent ensemble, les plus gros individus mangent en premier et peuvent attaquer les dragons plus petits qui tentent de se nourrir; si un petit animal meurt, il est habituellement cannibalisé. Si on applique ce type de comportement à des sites contenant de possibles groupes de dromaeosaures ayant pu chasser en meute, il semble tout à fait comparable au mode d'alimentation des dragons de Komodo ou des crocodiles. Les squelettes de Deinonychus découverts sur ces sites sont de jeunes adultes, avec des pièces manquantes qui peuvent avoir été mangées par d'autres Deinonychus, ce qu'une étude de Roach présente comme une preuve que ces animaux ne coopéraient pas à la chasse[32] .
En 2007, les scientifiques ont décrit la première série d'empreintes de pas d'un grand dromaeosaure, dans le Shandong, en Chine. En plus de confirmer l'hypothèse selon laquelle la griffe en faucille ne touchait pas le sol, les empreintes de pas laissées par des individus de la taille d'Achillobator montraient des traces de six individus d'environ la même taille marchant le long d'une côte. Les individus étaient espacés d'environ un mètre et se déplaçaient dans le même sens à un rythme assez lent. Les auteurs du document décrivent ces empreintes comme la preuve que certaines espèces de dromaeosaures vivaient en groupe. Alors que ces empreintes ne représentent manifestement pas le comportement d'un groupe en train de chasser, elles ne permettent pas d'éliminer l'idée que des dromaeosaures peuvent avoir chassé en bandes[7].
Vol battu et vol plané
 Fossile de Microraptor gui avec ses plumes alaires.
Fossile de Microraptor gui avec ses plumes alaires.On pense qu'au moins deux espèces de dromaeosauridés ont été capables de voler ou de planer. La première, Rahonavis ostromi (initialement classée dans les oiseaux mais qui s'est révélée être un dromaeosauridé dans des études ultérieures[5],[33]) aurait pu avoir été capable de voler comme le montrent ses longs membres antérieurs qui portent à l'évidence des excroissances osseuses servant de points d'attaches à de longues plumes robustes[34]. Les membres antérieurs des Rahonavis étaient plus puissamment bâtis que ceux de l'archéoptéryx et montrent à l'évidence des points d'insertions pour les puissants ligaments nécessaires au vol battu. Luis Chiappe en a conclu que ces adaptations pouvaient probablement permettre à Rahonavis de voler, mais qu'il devait être plus maladroit dans les airs que les oiseaux de nos jours[35].
Une autre espèce de dromaeosauridé, Microraptor gui, a dû être capable de planer à l'aide de ses ailes bien développées sur les avants et arrières. Une étude réalisée en 2005 par Sankar Chatterjee a suggéré que les ailes de Microraptor devaient fonctionner comme celles d'un biplan avec ses deux paires décalées et qu'il avait vraisemblablement utilisé un style phygoïde de glisse, se lançant d'un coup dans les airs à partir d'un arbre et décrivant une trajectoire en forme de « U » puis arriver sur un autre arbre, la queue et les ailes postérieures aidant à contrôler la position et la vitesse. Chatterjee a également constaté que Microraptor avait les moyens de base pour le vol battu en plus du vol plané[10].
Le vol battu a également été suggéré comme possible pour Cryptovolans pauli (dont le nom de genre signifie « voleur caché ») mais Cryptovolans est probablement un synonyme de Microraptor[36],[37].
Sens
La comparaisons des anneaux sclérotiques de plusieurs dromæosauridés (Microraptor, Sinornithosaurus et Velociraptor) à ceux des oiseaux et reptiles actuels montre que certains dromæosauridés (comme Microraptor et Velociraptor) peuvent avoir été des prédateurs nocturnes, tandis que Sinornithosaurus aurait pu être un chasseur cathéméral (actif jour et nuit pendant de courtes périodes)[38].
Les études des bulbes olfactifs des dromæosauridés ont révélé que ces bulbres étaient proportionnellement de la même taille que ceux d'autres théropodes non-aviaires et des oiseaux actuels avec un sens de l'odorat aussi développé que celui des tyrannosauridés et de l'Urubu à tête rouge ce qui reflète probablement l'importance de l'odorat dans l'activité quotidienne des dromæosauridés pour trouver de la nourriture[39],[40].
Classification
Relations avec les oiseaux
Articles détaillés : Histoire évolutive des oiseaux et Dinosaures à plumes.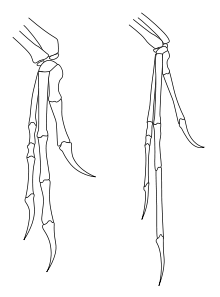 Comparaison des membres antérieurs de Deinonychus (à gauche) et d'Archaeopteryx (à droite), une des parties du squelette ayant le plus de similitude.
Comparaison des membres antérieurs de Deinonychus (à gauche) et d'Archaeopteryx (à droite), une des parties du squelette ayant le plus de similitude.Les Dromaeosauridae partagent de nombreuses caractéristiques avec les oiseaux primitifs (clade des Avialae ou des Aves). La nature exacte de leur relation avec les oiseaux a fait l'objet d'un grand nombre d'études et les hypothèses sur cette relation ont beaucoup changé en fonction des nouveaux éléments de preuve disponibles. Jusqu'en 2001, Mark Norell qui avait mené une vaste étude sur les coelurosauriens en était arrivé au résultat provisoire que les dromæosauridés étaient les dinosaures les plus étroitement liés aux oiseaux, les troodontidés étant un exogroupe plus lointain. Il avait même suggéré que Dromaeosauridae pourrait être paraphylétique à Avialae[41]. En 2002, Hwang et ses collègues ont utilisé les travaux de Norell, y ont ajouté de nouvelles découvertes sur ces fossiles, pour en conclure que les oiseaux (Avialae) devaient être considérés comme des cousins des dromæosauridés et des troodontidés[11].
Le consensus actuel est en accord avec les résultats de Hwang (2002); les dromæosauridés sont les plus proches parents des troodontidés et forment avec eux le clade Deinonychosauria. Les deinonychosauriens sont à leur tour le taxon frère des Avialae et en sont donc les plus proches parents[42]. Les paléontologues ont conclu qu'il n'y avait pas encore assez de preuves pour savoir si des dromaeosaures ont été les premiers vertébrés à pouvoir voler ou planer, ou s'ils descendent d'ancêtres qui pouvaient le faire[43].
Théories alternatives et impossibilité de voler
Les dromæosauridés sont si proches des oiseaux actuels qu'ils ont conduit certains chercheurs à soutenir qu'ils devraient être classés dans les oiseaux. Tout d'abord, du fait qu'ils avaient des plumes, les dromaeosaures (avec de nombreux autres coelurosauriens théropodes) sont des « oiseaux » suivant la définition traditionnelle du mot « oiseau » ou « Aves », qui est fondée sur la possession de plumes. Toutefois, d'autres scientifiques, comme Lawrence Witmer, ont fait valoir qu'appeler un théropode comme Caudipteryx un oiseau parce qu'il avait des plumes peut étendre tellement le sens du mot qu'il n'aurait plus aucun sens utile[44] (Voir l'article sur les Avialae pour une discussion sur les définitions des clades Aves et Avialae.).
 Moulage d'un fossile à plumes de microraptorien NGMC 91, n'ayant pas reçu de nom à ce jour
Moulage d'un fossile à plumes de microraptorien NGMC 91, n'ayant pas reçu de nom à ce jourAu moins deux écoles de chercheurs proposent que les dromaeosaures descendent d'ancêtres volants. L'hypothèse selon laquelle les dromaeosaures ont eu un ancêtre volant est parfois appelés Birds Came First (BCF) (Les oiseaux sont apparus d'abord). George Olshevsky est généralement crédité comme le premier auteur de cette théorie[45]. De son côté, Gregory S. Paul a fait remarquer que de nombreuses caractéristiques du squelette des dromaeosauridés pouvaient être interprétées comme une preuve que l'ensemble du groupe avait évolué à partir d'ancêtres dinosauriens volants, peut-être quelque chose comme Archaeopteryx. Dans ce cas, les plus grands dromæosauridés auraient été secondairement incapables de voler, un peu comme l'autruche actuelle[19]. En 1988, Paul a suggéré que les dromæosauridés pouvaient effectivement être plus étroitement apparentés aux oiseaux modernes que l’Archaeopteryx. Cependant, en 2002, Paul a placé les dromaeosaures et les Archaeopteryx comme les plus proches parents les uns des autres[46].
En 2002, Hwang a constaté que Microraptor était le dromaeosaure le plus primitif[11]. Xu et ses collaborateurs en 2003 a utilisé la position basale de Microraptor, ainsi que sa possession de plumes et d'ailes, comme une preuve que les ancêtres des dromaeosauridés pouvaient planer. Si cela est vrai, les plus grands des dromaeosaures seraient secondairement devenus terrestres ayant perdu la capacité de planer au cours de leur histoire évolutive[16].
Toujours en 2002, Steven Czerkas décrit Cryptovolans, peut-être un synonyme de Microraptor. Il a reconstruit le fossile de façon inexacte en lui donnant seulement deux ailes et il a donc soutenu que les dromaeosaures étaient des animaux capables de voler, plutôt que de se laisser planer. Par la suite, il en a publié une reconstruction révisée qui est correspond à celle d'un Microraptor[36].
D'autres chercheurs, comme Larry Martin croient que les dromaeosaures, ainsi que tous les maniraptoriens ne sont pas des dinosaures. Martin a affirmé pendant des décennies que les oiseaux n'étaient pas apparentés aux maniraptoriens, mais en 2004 il a changé de position et il admet maintenant que les deux sont les plus proches parents de la même famille. Martin croit que maniraptoriens sont devenus des oiseaux incapables de voler et que les oiseaux dérivent d'archosaures non-dinosauriens de sorte que la plupart des espèces anciennement appelées théropodes ne devraient plus maintenant être classées dans les dinosaures[47]
 Ce spécimen d’Archaeopteryx montre qu'il avait lui aussi un deuxième orteil hyperrétractile[48]
Ce spécimen d’Archaeopteryx montre qu'il avait lui aussi un deuxième orteil hyperrétractile[48].
En 2005, Mayr et Peters ont décrit l'anatomie d'un spécimen très bien conservé d'Archaeopteryx et ont pu constater que son anatomie était plus proche des théropodes non-aviaires qu'on ne le pensait. Plus précisément, ils ont trouvé que l'archéoptéryx avait un palatin primitif, un hallux non inversé et un deuxième orteil rétractile. Leur analyse phylogénétique est arrivé au résultat controversé que Confuciusornis était plus proche de Microraptor que d’Archaeopteryx, faisant des Avialae un taxon paraphylétique. Ils ont également suggéré que les paraviens ancestraux avaient été en mesure de voler ou de planer et que les dromaeosaures et les troodontidés sont devenus secondairement incapables de voler (ou ont perdu leur capacité de planer)[49],[50]. Corfe et Butler ont critiqué ce travail pour des raisons méthodologiques[51].
Un défi pour l'ensemble de ces hypothèses est venu quand Turner et ses collègues ont décrit en 2007 une nouveau dromaeosauridé, Mahakala, qu'ils ont estimé être le membre le plus basal et la plus primitif des Dromaeosauridae, encore plus primitif que Microraptor. Mahakala avait des bras courts et n'avait pas la possibilité de planer. Turner en a également déduit que le vol n'avait pu apparaitre que chez les Avialae, et ces deux points feraient que les Dromaeosauridé primitifs ne pouvaient pas planer ou voler. Sur la base de cette analyse cladistique, Mahakala suggère que les ancêtres des dromæosauridés ne volaient pas[52].
Taxonomie
La paternité de la famille des Dromaeosauridae est attribuée à William Diller Matthew et Barnum Brown, qui ont créé la sous-famille des Dromaeosaurinae de la famille des Deinodontidae en 1922. Cette sous-famille ne contenait que le nouveau genre Dromaeosaurus[53]. Les Dromaeosauridae avec les Troodontidae ont formé l'infra-ordre des Deinonychosauria.
Les sous-familles de Dromaeosauridae changent fréquemment de contenu du fait de nouvelles découvertes, mais sont formées généralement des groupes ci-dessous. Un certain nombre de dromæosauridés n'ont pas été affectés à une sous-famille particulière, souvent parce qu'ils sont trop mal conservés pour être placés avec certitude dans l'analyse phylogénétique (voir la section Phylogénie ci-dessous), ou parce qu'ils sont trop primitifs pour les subdivisions primaires des Dromaeosauridae (Mahakala, par exemple, qui est le plus primitif de tous les dromaeosauridés connus est placé en dehors de toutsous-groupe). La sous-famille la plus basale de dromæosauridés se trouve souvent être les Unenlagiinae[15]. Ce groupe mal connu est la sous-famille la plus mal étudiée de dromaeosauridés et il est possible que certains ou la totalité de ses membres appartiennent à d'autres familles que celle des Dromaeosauridae[54]. Les plus grands, les membres vivant sur le sol comme Buitreraptor et Unenlagia montrent d'importantes adaptations au vol, bien qu'ils étaient probablement trop grands pour décoller. Toutefois un membre de ce groupe, Rahonavis, est plus petit, avec des ailes bien développées présentant des signes de points d'insertions de ligaments pour les plumes de vol et il est très probable qu'il pouvait voler. Le clade suivant le plus primitif de dromaeosauridés est le Microraptoria. Ce groupe comprend plusieurs des plus petits dromaeosaures, qui montrent des adaptations de vie arboricole. Toules les traces de peau connues de dromaeosauridés sont originaires de ce groupe et tous montrent une importante couverture de plumes et des ailes bien développées. Comme les unenlagiinés, certaines espèces peuvent avoir été capables de voler de façon active. La sous-famille des Velociraptorinae inclut traditionnellement Velociraptor, Deinonychus et Saurornitholestes et, alors que la découverte de Tsaagan vient renforcer ce groupe, l'inclusion de Saurornitholestes est encore incertaine. Les Dromaeosaurinae consistent généralement en espèce de taille moyenne à grande, avec des têtes généralement carrées (alors que les autres sous-familles ont généralement un museau étroit).
 Mahakala, le plus primitif des dromaeosauridés connus
Mahakala, le plus primitif des dromaeosauridés connus Austroraptor, un unenlagiiné
Austroraptor, un unenlagiiné Sinornithosaurus, un microraptoriné
Sinornithosaurus, un microraptoriné Saurornitholestes, un eudromaeosaurien
Saurornitholestes, un eudromaeosaurienLa classification suivante des différents genres de dromæosauridés suit le tableau fourni par Holtz en 2010, sauf indication contraire[55].
- Famile Dromaeosauridae
- Dromaeosauroides
- Luanchuanraptor
- Mahakala
- Pamparaptor[56]
- Ornithodesmus
- Tianyuraptor
- Variraptor (=Pyroraptor?)
- Sous-famille Microraptorinae
- Graciliraptor
- Hesperonychus
- Microraptor (=Cryptovolans?)
- Sinornithosaurus
- Sous-famille Unenlagiinae[57]
- Austroraptor[58]
- Buitreraptor
- Neuquenraptor
- Rahonavis
- Shanag?
- Unenlagia
- Unquillosaurus
- Clade Eudromaeosauria[59]
- Sous-famille Dromaeosaurinae
- Sous-famille Saurornitholestinae
- Sous-famille Velociraptorinae
Phylogénie
Les Dromaeosauridae ont d'abord été définis comme un clade par Paul Sereno en 1998, formant le groupe naturel le plus inclusif contenant Dromaeosaurus mais pas Troodon, Ornithomimus ou Passer. Les différentes sous-familles ont également été redéfinies comme clades, généralement définie comme toutes les espèces plus proche de l'homonyme du groupe que de Dromaeosaurus ou de tout homonyme des autres sous-clades (par exemple, Makovicky a défini les Unenlagiinae comme le clade de tous dromæosauridés plus proches de Unenlagia que de Velociraptor). Le groupe Microraptoria est le seul sous-clade des dromaeosauridés qui n'ait pas été converti à partir d'une sous-famille. Senter et ses collègues ont expressément utilisé un nom de la sous-famille sans le suffixe -inae de la classification linnéenne pour éviter les problèmes rencontrés avec la construction d'un taxon traditionnel du groupe familial, le groupe pouvant un jour être placé en dehors des Dromaeosauridae[42]. Sereno a proposé une définition révisée du sous-groupe contenant Microraptor afin de s'assurer qu'il relèverait bien des Dromaeosauridae, et a créé la sous-famille des Microraptorinae, en l'imputant à Senter, bien que cette utilisation ne soit apparue que sur sa base de données en ligne et n'ait pas été officiellement publiée[60].
Le cladogramme ci-dessous résulte d'une étude en 2009 par les paléontologues Nicholas Longrich et Philip J. Currie[59].
Dromaeosauridae Mahakala
Unenlagiinae Rahonavis
unnamed Unenlagia
Unenlagia? paynemili
Neuquenraptor
Buitreraptor
unnamed Microraptorinae Shanag
Graciliraptor
Sinornithosaurus
Hesperonychus
Eudromaeosauria Saurornitholestinae Saurornitholestes
unnamed unnamed Velociraptorinae unnamed unnamed Tsaagan
Dromaeosaurinae .
Caractéristiques
 Fossile de dromaeosauridé exposé au Hong Kong Science Museum.
Fossile de dromaeosauridé exposé au Hong Kong Science Museum.Les dromaeosauridés sont définis par les caractères suivants :
- de courts os frontaux en forme de T qui forment la limite antérieure de la fenêtre supratemporale,
- un plateau postérolatéral au niveau du squamosal,
- un processus latéral de l'os carré qui vient en contact avec le quadratojugal,
- des parapophyses surélevées, pétiolées au niveau des vertèbres dorsales,
- une modification de la griffe du deuxième orteil,
- des chevrons et des prézygapophyses allongées s'étendant sur plusieurs vertèbres au niveau des vertèbres caudales,
- la présence d'une fossette subglénoïde sur la coracoïde[6].
Les dromaesauridés dans la culture populaire
Un Deinonychus figure dans le roman de John Brosnan paru en 1984 Carnosaur et son adaptation cinématographique a peu retenu l'attention des médias[61]. Velociraptor, un dromaeosauridé, a beaucoup plus attiré l'attention sur lui après avoir figuré en bonne place en 1993 dans le film de Steven Spielberg Jurassic Park. Toutefois, les dimensions du Velociraptor du film sont beaucoup plus grandes que celles des membres les plus importants de ce genre. Robert Bakker a rappelé que Spielberg avait été déçu par les dimensions du Velociraptor et qu'il l'avait grandi, ajoutant peu de temps après qu'il l'avait nommé Utahraptor un animal qui était de taille plus comparable voire plus grand[62]. Gregory S. Paul, dans son livre Predatory Dinosaurs of the World, considére Deinonychus antirrhopus comme une espèce de Velociraptor et le rebaptise Velociraptor antirrhopus[46]. Ce point de vue n'a pas été beaucoup partagé[6],[63],[64]. Michael Crichton a adopté la synonymisation de ces deux genres dans ses romans, qui ont inspiré les deux premiers films. La représentation des dromaeosauridés dans le film Jurassic Park, alors qu'ils étaient tout juste connus à cette époque, est inexacte à bien des égards, notamment l'absence de plumes. Bien que Jurassic Park III ait tenté de corriger cette erreur en ajoutant des pièces ressemblant à des plumes autour de la tête de certains de ses dromaeosaures, ces dernières ne ressemblent pas par leur aspect ou leur répartition aux vraies plumes retrouvées sur les fossiles de dromaeosauridés.
Les dromæosauridés apparaissent également dans bon nombre de films Le Petit Dinosaure, à partir du troisième épisode.
Des dromæosauridés apparaissent dans la série Nick Cutter et les Portes du Temps/Les Portes du Temps, ou Primeval en version originale à partir de la saison 2, dans la série, ils sont représentés portant des protoplumes qu'ils utilisent pour intimider leurs ennemis. Bien qu'au départ identifiés comme des Deinonychus, à partir de la saison 4, le site internet de la série les mentionnent comme des Dromaeosaurus bien qu'ils ont le crâne et une taille comparable à celle de Deinonychus.
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Dromaeosauridae » (voir la liste des auteurs)
- Case, J.A., Martin, J.E., and Reguero, M. (2007). "A dromaeosaur from the Maastrichtian of James Ross Island and the Late Cretaceous Antarctic dinosaur fauna." Pp. 1–4 in Cooper, A., Raymond, C., and Team, I.E. (eds.), Antarctica: a Keystone in a Changing World – Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences, U.S. Geological Survey Open-File Report 2007-1047, SRP 083. U.S. Geological Survey, Washington, D.C.
- Metcalf, S.J., Vaughan, R.F., Benton, M.J., Cole, J., Simms, M.J. and Dartnall, D.L., « A new Bathonian (Middle Jurassic) microvertebrate site, within the Chipping Norton Limestone Formation at Hornsleaslow Quarry, Gloucestershire », dans Proceedings of the Geologists’ Association, vol. 103, 1992, p. 321–342 [lien DOI]
- J. H. Ostrom, « Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana », dans Peabody Museum of Natural History Bulletin, vol. 30, 1969, p. 1–165 [texte intégral [PDF]]
- Paul, Gregory S. (2006) "Drawing Dinosaurs"(2006). "Geotimes", January 2006. http://www.geotimes.org/jan06/feature_drawingdinos.html
- Peter J. Makovicky, « The earliest dromaeosaurid theropod from South America », dans Nature, vol. 437, no 7061, 2005, p. 1007–1011 [lien PMID, lien DOI]
- (en) Norell, M.A., and Makovicky, P.J., The Dinosauria, Berkeley, University of California Press, 2004 (ISBN 0-520-24209-2), « Dromaeosauridae », p. 196–210
- Rihui Li, « Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China », dans Naturwissenschaften, vol. 95, no 3, 2007, p. 185–91 [texte intégral, lien PMID, lien DOI]
- Csiki Z., « An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania », dans Proceedings of the National Academy of Sciences of the United States of America, vol. preprint, no 35, in press, p. 15357–61 [lien PMID, lien DOI] Supporting Information
- Mark A. Norell, « Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis », dans American Museum Novitates, vol. 3282, 1999, p. 1–45 [texte intégral]
- S. Chatterjee, « Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui », dans Proceedings of the National Academy of Sciences, vol. 104, no 5, 2007, p. 1576–1580 [texte intégral, lien PMID, lien DOI]
- S.H. Hwang, « New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China », dans American Museum Novitates, no 3381, 2002, p. 44pp [texte intégral]
- A. Perle, « A new maniraptoran theropod – Achillobator giganticus (Dromaeosauridae) – from the Upper Cretaceous of Burkhant, Mongolia », dans Contributions of the Mongolian-American Paleontological Project, vol. 101, 1999, p. 1–105
- Britt, « New osteological data and the affinities of Utahraptor from the Cedar Mountain Fm. (Early Cretaceous) of Utah », dans Journal of Vertebrate Paleontology, vol. 21, no 3, 2001, p. 36A [lien DOI]
- Naish, D. Hutt, and Martill, D.M. (2001). "Saurischian dinosaurs: theropods." in Martill, D.M. and Naish, D. (eds). Dinosaurs of the Isle of Wight. The Palaeontological Association, Field Guides to Fossils. 10, 242–309.
- A.S. Turner, « A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia », dans American Museum Novitates, vol. 3557, 2007, p. 1–27 [texte intégral [PDF], lien DOI (pages consultées le 29 mars 2007)]
- Xu Xing, « Four-winged dinosaurs from China », dans Nature, vol. 421, no 6921, 2003, p. 335–340 [lien PMID, lien DOI]
- Xu, X., Wang, X.-L., and Wu, X.-C., « A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China », dans Nature, vol. 401, no 6750, 1999, p. 262–266 [lien DOI]
- A.H. Turner, « Feather quill knobs in the dinosaur Velociraptor », dans Science, vol. 317, no 5845, 2007, p. 1721 [texte intégral [PDF], lien PMID, lien DOI]
- Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 472 pp.
- R. Prum, « The evolutionary origin and diversification of feathers », dans The Quarterly Review of Biology, vol. 77, no 3, 2002, p. 261–295 [lien PMID, lien DOI]
- Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pg 1–6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- Manning, P.L., Payne, D., Pennicott, J., Barrett, P.M., and Ennos, R.A., « Dinosaur killer claws or climbing crampons? », dans Biology Letters, vol. 2, no 1, 2005, p. 110–112 [texte intégral, lien PMID, lien DOI]
- Davies, S.J.J.F. (2002) "Ratites and Tinamous" Oxford University Press. New York, USA
- Gilliard, E. T. (1958) "Living birds of the world" Doubleday, garden City, NY. USA. 400p.
- Christopher P. Kofron, « Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia », dans Journal of Zoology, vol. 249, 1999, p. 375–381 [lien DOI]
- Christopher P. Kofron, « Case histories of attacks by the southern cassowary in Queensland », dans Memoirs of the Queensland Museum, vol. 49, no 1, 2003, p. 335–338
- Kent H. Redford, « Notes on the biology and song of the red-legged seriema (cariama cristata) », dans Journal of Field Ornithology, vol. 57, no 4, 1986, p. 261–269 [texte intégral]
- P.L. Manning, « Biomechanics of dromaeosaurid dinosaur claws: Application of X-ray microtomography, nanoindentation, and finite element analysis », dans The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, vol. 292, no 9, 2009, p. 1397–1405 [lien DOI]
- « Velociraptor's 'killing' claws were for climbing », dans New Scientist, no 2725, septembre 2009 [texte intégral (page consultée le 2009-09-15)]
- Senter, P., « Pedal function in deinonychosaurs (Dinosauria: Theropoda): a comparative study », dans Bulletin of the Gunma Museum of Natural History, vol. 13, 2009, p. 1–14
- W. D. Maxwell, « Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations », dans Journal of Vertebrate Paleontology, vol. 15, no 4, 1995, p. 707–712 [lien DOI]
- B. T. Roach, « A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs », dans Bulletin of the Peabody Museum of Natural History, vol. 48, no 1, 2007, p. 103–138 [lien DOI]
- M.A. Norell, « A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia) », dans American Museum Novitates, vol. 3545, 2006, p. 1–51 [lien DOI]
- Catherine A. Forster, « The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar », dans Science, vol. 279, no 5358, 1998a, p. 1915–1919 [lien PMID, lien DOI]. (HTML abstract).
- (en) L.M. Chiappe, Glorified Dinosaurs: The Origin and Early Evolution of Birds, Sydney, UNSW Press, 2007 (ISBN 978-0-471-24723-4)
- Czerkas, S.A., Zhang, D., Li, J., and Li, Y. (2002). "Flying Dromaeosaurs", in Czerkas, S.J. (ed.): Feathered Dinosaurs and the Origin of Flight: The Dinosaur Museum Journal 1. Blanding: The Dinosaur Museum, 16–26.
- Alan Feduccia, « Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence », dans Journal of Morphology, vol. 266, no 2, 2005, p. 125–166 [lien PMID, lien DOI]
- Schmitz, L.; Motani, R., « Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology », dans Science, vol. in press, 2011 [lien DOI]
- Zelenitsky, D.K.; Therrien, F.; Kobayashi, Y., « Olfactory acuity in theropods: palaeobiological and evolutionary implications », dans Proceedings of the Royal Society B, vol. 276, no 1657, 2009, p. 667–673 [lien DOI]
- Zelenitsky, D.K.; Therrien, F.; Ridgely, R.C.; McGee, A.R.; Witmer, L.M., « Evolution of olfaction in non-avian theropod dinosaurs and birds », dans Proceedings of the Royal Society B, vol. in press, 2011 [lien DOI]
- Norell, M. Clark, J.M., Makovicky, P.J. (2001). "Phylogenetic relationships among coelurosaurian theropods." New Perspectives on the Origin and Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom”, Yale Peabody Museum: 49–67
- Phil Senter, « Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda) », dans Bulletin of the Gunma Museum of Natural History, vol. 8, 2004, p. 1–20
- Kevin Padian, « Origin of flight: Could ‘four-winged’ dinosaurs fly? », dans Nature, vol. 438, no 7066, 2005, p. E3 [lien DOI]
- Witmer, L.M. (2005) “The Debate on Avian Ancestry; Phylogeny, Function and Fossils.” In “Mesozoic Birds: Above the Heads of Dinosaurs”, pp. 3–30. ISBN 0-520-20094-2
- Olshevsky, George. (1994). "The birds first? A theory to fit the facts — evolution of reptiles into birds". Omni, June 1994. Volume 16 No. 9
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster. 464 pp.
- L.D. Martin, « A basal archosaurian origin for birds », dans Acta Geologica Sinica, vol. 50, no 6, 2004, p. 978–990 [texte intégral [PDF]]
- National Geographic News- Earliest Bird Had Feet Like Dinosaur, Fossil Shows – Nicholas Bakalar, December 1, 2005, Page 2. Retrieved 2006-10-18.
- G Mayr, « A Well-Preserved Archaeopteryx Specimen with Theropod Features », dans Science, vol. 310, no 5753, 2005, p. 1483–1486 [lien PMID, lien DOI]
- G Mayr, « Response to comment on ‘A Well-Preserved Archaeopteryx Specimen with Theropod Features’ », dans Science, vol. 313, 2006, p. 1238c [lien DOI]
- IJ Corfe, « Comment on ‘A Well-Preserved Archaeopteryx Specimen with Theropod Features’ », dans Science, vol. 313, no 5791, 2006, p. 1238b [lien PMID, lien DOI]
- Alan H. Turner, « A basal dromaeosaurid and size evolution preceding avian flight », dans Science, vol. 317, no 5843, 2007, p. 1378–1381 [lien PMID, lien DOI]
- W. D. Matthew, « The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta », dans Bulletin of the American Museum of Natural History, vol. 46, 1922, p. 367–385 [texte intégral]
- Agnolin, F.L.; Novas, F.E., « Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)? », dans Anais da Academia Brasileira de Ciências, vol. 83, 2011, p. 117–162 [texte intégral [PDF], lien DOI (pages consultées le 23 avril 2011)]
- Holtz, Thomas R. Jr. (2010) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Juan D. Porfiri, « A new small deinonychosaur (Dinosauria: Theropoda) from the Late Cretaceous of Patagonia, Argentina », dans Anais da Academia Brasileira de Ciências, vol. 83, no 1, 2011, p. 109–116 (ISSN 0001-3765) [texte intégral]
- Bonaparte, (1999).
- Fernando E. Novas, « A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids », dans Proceedings of the Royal Society B: Biological Sciences, vol. 276, no 1659, 2009, p. 1101–7 [lien PMID, lien DOI]
- N.R. Longrich, « A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America », dans PNAS, vol. 106, no 13, 2009, p. 5002–7 [lien PMID, lien DOI]
- Sereno, P. C. 2005. Stem Archosauria—TaxonSearch (version 1.0), November 7, 2005
- Huett, Kim, You only Live Once, 2007
- (en) Robert T. Bakker, Raptor Red, New York, Bantam Books, 1995 (ISBN 0-553-57561-9), p. 4
- B.P. Pérez-Moreno, « A theropod dinosaur from the Lower Cretaceous of southern France », dans Dinosaurs and Other Fossil Reptiles of Europe, Second Georges Cuvier Symposium, Montbéliard; Revue de Paléobiologie, Volume spécial, vol. 7, 1994, p. 173–188
- P. J. Currie, « New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda) », dans Journal of Vertebrate Paleontology, vol. 15, no 3, 1995, p. 576–591 [lien DOI] (abstract)
Deinonychus (« griffe terrible ») et les autres dromæosauridés (Dromaeosauridae les « lézards qui courent ») étaient de petits prédateurs agressifs. Par leur « intelligence » ils surpassaient la plupart des théropodes et ils étaient formidablement armés. Les mâchoires des dromæosauridés étaient hérissées de grandes dents recourbées et coupantes comme des lames, les mains et les pieds portaient trois doigts munis de grosses griffes crochues. L'énorme griffe du deuxième orteil était deux fois plus longue que les autres. Les dromaeosauridés mesuraient entre 2 et 6 m environ. Nombre d'entre eux semblent aujourd'hui avoir été recouverts de plumes. Ils apparurent probablement au Jurassique, même si les fossiles retrouvés datent du Crétacé. Les dromæosauridés se répandirent largement sur les continents de l'hémisphère nord après leur séparation du Gondwana, le mégacontinent austral.
« Reptile agile », du grec dramaios (« agile à la course ») et sauros (« reptile »).
Le Dromaeosaurus est un petit dinosaure cœlurosaurien du Crétacé terminal.
Son poids peut être estimé à une quarantaine de kilos pour une longueur totale d'environ 2,50 à 3 mètres.
C'était un reptile bipède carnivore muni de dents petites et tranchantes ainsi que d'une griffe recourbée et acérée à l'extrémité du doigt interne de chaque pied.
Le cou de l'animal était plutôt long, mais étroit et puissant.
Le Dromaeosaurus avait un crâne large contenant un cerveau assez volumineux ainsi que de grandes cavités oculaires.
Le Dromaeosaurus a été identifié pour la première fois dans le Crétacé supérieur de l'Amérique du Nord (Alberta, Canada). Ce petit théropode est connu depuis peu en Provence par deux genres distincts :
- Le premier genre est représenté par des vertèbres et des os des membres découverts dans le gisement des Grands-Creux (Bouches-du-Rhône) et dans la région de Fox-Amphoux (Var), ainsi que de nombreuses dents isolées provenant du Rognacien inférieur des environs de Trets et de Vitrolles (Bouches-du-Rhône). Sur la base de ces vestiges E. Buffetaut et J. Le Lœuff ont érigé un nouveau genre et une nouvelle espèce : le Variraptor mechinorum (le « voleur du Var », « mechinorum » en hommage à P. et A. Méchin, découvreurs d'une grande partie du matériel osseux connu à l'heure actuelle).
- Plus récemment (en 2000) Ronan et Taquet ont décrit un nouveau genre de Dromaeosauridae, Pyroraptor olympius, d'après l'étude de plusieurs pièces osseuses (phalanges unguéales, métatarse, dents et ulna) découvertes dans les sédiments bégudiens du gisement de La Boucharde (sud-est de Trets, Bouches-du-Rhône) . Ces mêmes auteurs considèrent le Variraptor mechinorum comme nomen dubium.
Voir aussi
- Amazoraptor
 Portail de la paléontologie
Portail de la paléontologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail des dinosaures
Portail des dinosaures Portail de la zoologie
Portail de la zoologie
















Wikimedia Foundation. 2010.