- Trochilidae
-
 Colibris
Colibris
Peinture de Ernst Haeckel parue dans
Kunstformen der Natur de 1904Classification (COI) Règne Animalia Embranchement Chordata Sous-embr. Vertebrata Classe Aves Ordre Apodiformes Famille Trochilidae
Vigors, 1825 Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsLa famille des trochilidés (ou Trochilidae) comprend environ 330 espèces d'oiseaux couramment appelées colibris[1]. Ils sont également surnommés « oiseaux-mouches » à cause de leur petite taille et leurs battements d'ailes rapides. Ces oiseaux colorés se rencontrent exclusivement dans les Amériques et on en rencontre des espèces à presque toutes les altitudes, latitudes et biotopes. Ils sont particuliers à plusieurs égards, ils peuvent voler vers l'arrière ou sur place et effectuer des voltiges impressionnantes. Ils sont capables d'une « hibernation » nocturne et de voir les ultra-violets. En outre, ils ont un métabolisme extrêmement élevé, un rapport entre la masse corporelle et la masse du cerveau tout aussi élevé, une capacité à faire des vocalises complexes et une capacité d'apprentissage des chants comparable à celle des Psittacidae et des Passeriformes.
Sommaire
Noms désignant les représentants de la famille
Les oiseaux-mouches ont des noms normalisés évocateurs : colibri, bec-en-faucille, ermite, porte-lance, campyloptère, mango, coquette, émeraude, dryade, saphir, ariane, brillant, inca, héliange, érione, haut-de-chausses, porte-traîne, métallure, sylphe et loddigésie.
Le terme « colibri » s'est imposé dans bon nombre de langues, y compris le français où il peut prendre plusieurs sens, ce qui entraîne une certaine confusion. Colibri désigne ainsi l'ensemble des Trochilidae, mais aussi l'un de ses genres, Colibri, et il est utilisé dans les noms normalisés de beaucoup de ses espèces.
Les Cubains préfèrent appeler ces oiseaux zunzuncito, mot espagnol qui veut dire « petit zunzun ». Zunzun est une onomatopée imitant le bruit fait par le battement très rapide des ailes de l'oiseau-mouche. Le terme anglophone hummingbird est riche de sens, car il dérive du verbe to hum, qui peut se traduire par vrombir et chantonner.
Description
Morphologie
Ce sont des oiseaux petits à minuscules qui mesurent de 2 à 22 cm environ et jusqu'à 35 cm chez des espèces dont la queue porte de longues rectrices. Ils disposent de dix rectrices sauf chez le Loddigésie admirable qui n'en possède que quatre dont, pour le mâle adulte, deux portent une palette à leur extrémité[2]. Leurs ailes sont longues et étroites et les battements de celles-ci sont très rapides c'est-à-dire de 8 à 80 hertz. La longueur de leur queue est variable. Leur bec fin est de longueur extrêmement variable selon les espèces, des becs courts de 1,5 cm à très longs de 10 cm, de rectiligne à nettement courbé. Les pattes sont courtes et se terminent par des pieds minuscules et fragiles. Elles sont Apodiformes, c'est-à-dire qu'elles ne leur permettent pas de se poser au sol, mais uniquement sur une branche.
La couleur du plumage varie du brun au vert ou du rouge au noir. Elle est souvent similaire à celle des fleurs que les oiseaux visitent, par mimétisme. Ils ont la gorge ou le dessus de la tête vivement colorés. Certaines plumes possèdent des plaques iridescentes réfléchissant des couleurs métalliques différentes selon l'angle d'où provient la lumière. Leurs plumes squameuses, aux barbules hypertrophiées, forment un ensemble compact sur le corps. Elles ne sont pas retenues entre elles comme chez les autres oiseaux. Cette structure spéciale des plumes iridescentes, à la forme aplatie, renferme des particules d'air. Les oiseaux-mouches disposent de 1 000 plumes en comparaison des 25 000 des cygnes[3] soit 300 plumes par gramme contre 4 pour les cygnes.
Archilochus alexandri mâle femelle 

Le dimorphisme sexuel est important chez les Trochilinae, les mâles sont plus colorés et le mimétisme des femelles est plus performant[4]. Alors que les différences entre espèces sont souvent difficiles à distinguer à l'œil nu chez les femelles et les juvéniles, la détermination des mâles est facile grâce aux nettes différences spécifiques de coloration. Chez les femelles des espèces aux mâles colorés ou chez les taxons plus ternes (Phaethornithinae), la longueur et la forme du bec ainsi que leurs chants sont des critères déterminants pour l'identification.
Le plus petit oiseau-mouche vit à Cuba. Avec ses 2 grammes, les femelles de colibri d'Hélène dépassent à peine 2 cm de longueur et pondent les plus petits œufs au monde tandis que les plus grands sont les colibris géants avec leurs 20 grammes et 21 cm[5].
Alimentation
Les oiseaux-mouches sont principalement nectarivores, le nectar constituant 90 % du régime alimentaire[5], mais également insectivores. Les insectes qu'ils capturent en vol, qu'ils récoltent sur les feuilles ou encore qu'ils retirent des toiles d'araignée leur sont indispensables comme apport en protéines. Certaines espèces peuvent se nourrir de la sève qui coule des arbres après que les pics y ont creusé à la recherche d'insectes ou de sève. Ils consomment en moyenne la moitié de leur poids en sucre chaque jour. Ils se nourrissent de 5 à 8 fois par heure à raison de 30 à 60 secondes à chaque fois, pour 3,14 à 7,6 calories par jour[5], ce qui est énorme en comparaison de leur taille.
La superficie de leur territoire est déterminée par l'abondance des fleurs dont ils se nourrissent.
 Oiseau-mouche tirant la langue
Oiseau-mouche tirant la langue
Les oiseaux-mouches atteignent et absorbent le nectar des fleurs grâce à leur longue langue extensible séparée en deux parties à leur extrémité et garnie de petites soies et de nervures. Préalablement renfermée dans une structure particulière attachée à leur bec en forme d'étui, leur langue entre et sort, telle un ressort, un peu à la manière d'un pic. Leur langue atteint le nectar dans la corolle, puis celui-ci monte le long de celle-ci par capillarité[6]. Il est finalement aspiré lorsque la langue est de retour dans le bec.

Certaines fleurs comme les Ornithophilous et les becs des oiseaux-mouches qui les visitent auraient connu un processus de coévolution[7],[8]. En effet, les fleurs bénéficient du passage des oiseaux-mouches dans leur corolle pour déposer leur pollen sur la tête de l'oiseau-mouche qui la visite. Ces fleurs profitent donc de cet oiseau en accroissant la probabilité que ce pollen soit déposé dans une autre fleur de la même espèce et la féconde. L'évolution du système buccal des oiseaux-mouches, langue et bec, s'est faite en parallèle avec l'adaptation des fleurs[9].
D'autre part, pour attraper plus facilement les insectes, lorsqu'ils ouvrent leur bec, la base de celui-ci s'agrandit. Ceci est unique chez les oiseaux[10].
Comportement
Territorialité
Article détaillé : Territorialité des oiseaux. Combat de mâles
Combat de mâlesLa plupart des espèces sont très territoriales et solitaires. Certaines espèces osent même attaquer des faucons qui s'aventurent sur leur territoire[5]. Les acrobaties aériennes seraient surtout composées de gestes voulant démontrer leur agressivité. La majorité des comportements observés visent à protéger la source de nourriture qu'ils revendiquent. La faiblesse de l'apport nutritif de leur alimentation par rapport à l'effort fait pour l'obtenir serait une explication pour cette agressivité[11]. Aussi ils défendent âprement leurs ressources vis-à-vis de concurrents, souvent de la même espèce. Les espèces différentes cohabitantes dans une même zone ne se nourrissent pas en général des mêmes fleurs ou sur le même étage de la flore.
Certaines espèces vivent en bande lors de leur migration tout en étant territoriaux lorsqu'ils sont établis dans leur zone de résidence. Les mâles combattent également pour les femelles pendant la saison de reproduction.
Reproduction
 Colibri à queue large avec ses petits
Colibri à queue large avec ses petitsOn ignore la plupart des comportements reproductifs de chacune des espèces. On suppose que le comportement du colibri d'Anna pourrait correspondre à celui de plusieurs espèces d'Amérique du Nord. Il arrive que le mâle soit plus petit que la femelle. En général, une fois accouplée, la femelle construit le nid et élève les petits seule; le mâle cherche à séduire d'autres femelles.
Généralement deux œufs minuscules sont pondus dans un nid en forme de coupe, construit avec de la matière végétale retenue par des toiles d'araignées ou des soies d'insectes. Les œufs sont également très bien protégés dans un nid ressemblant aux nœuds d'une branche. Certains spécimens de Phaethornithinae pondent dans le nid d'autres femelles[4]. L'incubation dure en général 14 à 19 jours. Les oisillons naissent aveugles et nus. Plus exactement, on peut observer deux rangées de plumes minuscules situées sur le dos. Celles-ci vibrent à l'arrivée de la mère et avertissent les poussins du moment tant attendu de la becquée. Ils sont nourris principalement avec des insectes durant une période de 7 à 12 jours, jusqu'à ce qu'ils soient capables de contrôler leur température. La femelle les nourrit ensuite de 18 à 25 jours après qu'ils ont quitté le nid[4].
Communication
L'appareil vocal de l'oiseau-mouche est simple en comparaison de celui des oiseaux dits chanteurs. En dépit de cet appareil peu développé, on sait que certains oiseaux-mouches des tropiques émettent des chants élaborés. Ceux-ci sont inaudibles pour les humains en raison de leurs fréquences trop élevées pour notre appareil auditif. Le chant des femelles est différent de celui des mâles[4]. Avec les Psittacidae et certains Passeriformes, ils sont les seuls oiseaux à pouvoir apprendre un chant[12].
En général, les oiseaux-mouches produisent trois types de signaux sonores, un léger bourdonnement, un bruit de crécelle et un gazouillis. Le bourdonnement, produit par les ailes, provient de la friction de l'air à travers les plumes. Le plus mélodieux des oiseaux-mouches serait le mâle colibri à queue large qui, comme un virtuose, laisse entendre un son très musical produit par ses ailes, un peu à la manière d'un criquet. Les différents bourdonnements permettent de distinguer certaines espèces l'une de l'autre. Cette technique d'identification aide les observateurs à bien les identifier sur le terrain et ceci malgré la distance. Les bourdonnements sont plus fréquemment émis par les mâles et il est possible d'en prendre conscience lors des démonstrations aériennes et des voltiges.
Ils communiquent également en utilisant leur plumage. Ainsi, perchés sur une branche tout en s'assurant d'être bien vus par un oiseau-mouche de passage, ils déploient les plumes de la gorge ou du cou afin de lui adresser un message sans équivoque. Ils agissent aussi de la sorte en exhibant leur queue bien ouverte. Ceci est plus fréquent chez les femelles et les jeunes qui désirent mettre en évidence les pointes blanches des plumes caudales.
Les voltiges aériennes sont principalement de deux types. La première parade se caractérise par un léger va-et-vient d'avant en arrière faisant face à un autre individu. Dans cette mimique en forme d'un petit arc horizontal, les plumes de la queue et de la gorge sont bien déployées. La seconde voltige aérienne, souvent appelée «le pendule», la manifestation est d'une plus grande intensité. Elle consiste en un grand arc de cercle exécuté de haut en bas adoptant la forme d'un grand «U» d'où l'oiseau peut s'élever de 1 à 12 mètres pour ensuite rejoindre son perchoir. À ce moment, il est possible d'entendre un bourdonnement produit par les ailes et des cris grinçants. Cette dernière parade est propre au mâle qui témoignerait ainsi de son agressivité vis-à-vis d'un autre individu de même sexe ou de sexe opposé. Quant à la première, elle ne saurait être l'apanage d'un seul sexe et serait encore là pour témoigner des sentiments hostiles envers d'autres oiseaux-mouches. Les recherches récentes n'excluent pas que ces parades et ces hautes voltiges soient utilisées en d'autres circonstances pour communiquer leurs intentions amoureuses à un partenaire de sexe opposé.
On connaît également une autre parade réalisée par les deux opposants simultanément, qu'on appelle : « vol vertical », les individus volent de haut en bas sur une certaine hauteur en se faisant face à une distance de moins de 1 mètre l'un de l'autre. Cette dernière parade peut être accompagnée de gazouillis.
On reconnaît également un autre vol dit «horizontal» qui se fait sur une courte trajectoire horizontale et peut être accompagné de gazouillis ou de bourdonnements. Ce comportement serait également utilisé pour témoigner son agressivité vis-à-vis d'un autre oiseau-mouche et comme possible parade nuptiale.
Sens
Article détaillé : vision des oiseaux.Comme beaucoup d'oiseaux, les trochilidés ont peu ou pas de sens de l'odorat. Par contre, leur vue est performante. Des expériences ont montré qu'ils étaient particulièrement attirés par les fleurs rouges, comptant y trouver le plus de nectar possible[13]. Ils sont cependant capables de comprendre que ce n'est pas forcément le cas et de changer leur habitude.
Comme les autres oiseaux, ils sont capables de distinguer les ultra-violets, pour des longueurs d'onde de 325 à 360 nanomètres. Outre que cela leur permet vraisemblablement de mieux distinguer certaines fleurs, ceci permet aussi aux femelles de mieux choisir leur mâle[4].
Métabolisme
 Colibri de Costa mâle au repos
Colibri de Costa mâle au reposLe métabolisme de ces oiseaux est très rapide, même pour les oiseaux, qui possèdent un métabolisme plus rapide que celui des mammifères. Un oiseau-mouche peut digérer une mouche à fruit en 10 minutes[5]. Leurs ailes battent à des allures vertigineuses, le rythme cardiaque du colibri à gorge bleue a été mesuré à plus de 1 260 battements par minute[14]. Ils consomment leur propre poids en nourriture chaque jour et ils sont souvent à quelques heures de mourir de faim. De ce fait, ils passent 70% de leur temps perchés[4], économisant leur énergie.
Cependant, ils sont capables de ralentir leur métabolisme jusqu'à la léthargie, par exemple la nuit, lorsque la nourriture n'est que difficilement disponible. Leur température peut baisser de 30 °C à 20 °C[4]. La fréquence cardiaque passe approximativement à 50 battements par minute. Leur respiration se ralentit nettement. Ils sont alors vulnérables car ils ne peuvent pas reprendre leur activité immédiatement[4].
Les oiseaux-mouches effectuant de longues migrations, comme le colibri à gorge rubis qui doit parcourir 800 km, doivent accumuler près de 40 à 50 % de leur poids en graisse pour pouvoir effectuer le voyage, cette graisse les ralentit considérablement[15].
Anatomie
Article détaillé : Anatomie des oiseaux.En comparaison avec leur poids, l'oiseau-mouche possède le plus gros cœur de tous les oiseaux. Ce dernier représente 2,4 % de sa masse corporelle contre 1 % chez le corbeau. Les performances de vol sont rendues possibles grâce à la masse musculaire du muscle pectoral qui constitue de 25 à 30 % de son poids en comparaison avec seulement 5 % chez l'humain. Chez les oiseaux-mouches, la souplesse de l'articulation de l'épaule les rend aptes à battre les ailes dans toutes les directions. En fait leur aile est constituée essentiellement de leur « main » alors que leurs bras sont très courts[16], contrairement aux autres oiseaux.
En outre, le sang des oiseaux-mouches a la plus haute teneur en hématie de tous les vertébrés, cela permet au sang de transporter une grande quantité d'oxygène aux muscles lors des efforts. Le volume d'oxygène brassé à chaque respiration est le double de celui d'un mammifère de même taille[17], en outre le système respiratoire des oiseaux est surement le plus performant de tous les vertébrés. De plus, la fréquence est d'environ 300 respirations par minute et de 500 en vol[17] contre 14 à 18 pour l'homme ou 30 pour les pigeons. Ainsi la densité en globules rouges est environ 6 590 000 unité par millimètre cube pour le Colibri rubis-topaze.
Si la plupart des oiseaux sont uricotéliques, excrétant de l’acide urique et des urates et non pas amino-uréolétique (excrétion d'une combinaison d’ammoniaque et d’urée), les Trochilidae, le sont partiellement[18], spécialement pour les petites espèces[19].
Une partie de la langue est cartilagineuse[6].
Locomotion

 Un oiseau-mouche (à gauche) et un sphingidé, le Moro-sphinx, (à droite) ont en commun une longue langue adaptée pour puiser le nectar et une capacité au vol stationnaire.
Un oiseau-mouche (à gauche) et un sphingidé, le Moro-sphinx, (à droite) ont en commun une longue langue adaptée pour puiser le nectar et une capacité au vol stationnaire.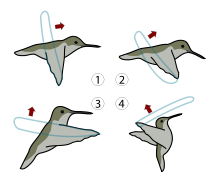 4 types de mouvement d'aile pour quatre directions
4 types de mouvement d'aile pour quatre directions Vol complet, vu de profil
Vol complet, vu de profilLes oiseaux-mouches sont capables de battre des ailes très rapidement, 8 à 10 Hz pour le colibri géant, 20 à 25 battements pour les espèces de taille moyenne et plus de 70 pour les plus petites. Leurs battements d'ailes produisent un vrombissement caractéristique.
La structure osseuse de leurs ailes que l'on ne retrouve qu'au niveau du poignet des autres oiseaux assure une mobilité des épaules unique alors qu'elle est axée sur le seul mouvement vertical dirigé de haut en bas pour les autres oiseaux. Les oiseaux-mouches, grâce aux particularités de leur squelette peuvent battre des ailes dans toutes les directions et faire des prouesses aériennes inégalées. Ils peuvent voler sur place en effectuant des mouvements de leurs ailes en forme de 8. Ils sont les seuls oiseaux capables de voler en arrière[21]. Leurs cabrioles peuvent être effectuées à une vitesse moyenne de 30 à 45 kilomètres par heure. Le vol d'avant en arrière ou vol en piqué peuvent atteindre 96 km par heure avec jusqu'à 200 battements par seconde pour un vol en plongée. Selon des chercheurs américains, cet oiseau parvient à faire du sur-place en battant des ailes à 75 % vers le bas et à 25 % vers le haut[22]. Cette façon de voler rappelle le vol de certains insectes comme celui des Sphingidae.
Chorologie
Écologie
Les oiseaux-mouches vivent dans des milieux très diversifiés, depuis les forêts andines, aux forêts tropicales en passant par les forêts tempérées d'arbres à feuilles caduques de la Terre de Feu ou d'Alaska. Certains oiseaux-mouches ne vivent que dans un biotope très particulier, voire un étage particulier d'une forêt, les autres étages étant occupés par d'autres espèces d'oiseaux-mouches. Ceci est lié à la symbiose évolutive entre certaines plantes et ces oiseaux[23]. Les oiseaux-mouches pollinisent 58 espèces de fleurs au Brésil. Les ananas sont pollinisés par les oiseaux mouches, il est d'ailleurs interdit d'importer ces oiseaux dans plusieurs régions du monde où ces pollinisateurs n’existent pas, comme aux îles Hawaii, pour ne pas menacer les rendements. Pour certaines espèces, ils sont même les seuls à pouvoir le faire[4] et sont donc vitaux pour la survie de ces plantes.
Certaines espèces d'Acari se servent des oiseaux-mouches comme espèces phorésiques. Comme eux, elles vivent du nectar et du pollen des fleurs. Elles s'abritent dans les narines des oiseaux quand ceux-ci visitent les fleurs et s'y reproduisent. Ces acariens ne nuisent pas aux oiseaux mais modifient indirectement leur alimentation et donc leur comportement[24],[25]. Au moins quarante espèces d'acariens de ce « type » ont été décrites[26].
Répartition
Ils peuplent la majeure partie des Amériques, à l'exception de la plus grande partie du Canada, mais se rencontrent cependant en Alaska. Ils sont particulièrement abondants dans la zone néotropicale. Les oiseaux-mouches habitent une large gamme de milieux, partout où poussent des plantes qui peuvent les alimenter en nectar, depuis le niveau de la mer jusqu'à près de 5 000 m d'altitude, avec la plus grande diversité en régions sub-montagneuses[4].
Le nombre de variété est la plus élevée en Équateur où on peut en observer 163 espèces. 31 espèces sont recensées en Guyane. En fait, 50 % des espèces vivent de part et d'autre de l'équateur dans une zone de 10º de latitude nord à 25° sud[4]. Leur distribution diminue au-delà de cette zone de manière régulière. Ainsi seulement 51 espèces sont recensées au Mexique et 21 au-delà. 16 s'y reproduisent, 1 en est un migrateur régulier, tandis que les 4 autres sont considérées comme des visiteurs occasionnels. Parmi celles-ci, 4 d'entre-elles se retrouvent au Canada dont le colibri roux, qui se rencontre jusqu'aux côtes sud de l'Alaska.
La distribution des espèces au sud est plus faible encore puisqu'il n'existe que 20 espèces au-dessous de 20º de latitude. Seul le colibri à dos vert traverse le détroit de Magellan pour nicher en Terre de Feu.
19 ont été comptabilisées aux Antilles et 2 dans l'archipel Juan Fernández.
Migration
Article détaillé : migration des oiseaux.La plupart des oiseaux-mouches ne migrent pas. Un certain nombre d'espèces effectue des migrations altitudinales saisonnières. C'est le cas notamment de certains colibris des régions montagneuses du Costa Rica qui doivent se replier pendant la saison des pluies vers des zones de plus basse altitude, pour pallier la raréfaction des fleurs et donc du nectar. Seules demeurent sur place les espèces les plus opportunistes capables de s'adapter à d'autres sources de nourriture.
Une minorité d'espèces effectue des migrations sur de longues distances. Les vols migratoires s'effectuent presque toujours au-dessus de la terre ferme. Ils sont entrecoupés de haltes plus ou moins longues qui dépendent notamment de l'abondance des sources de nourriture. Ainsi, sept espèces de colibris nichant dans la moitié occidentale des États-Unis migrent vers le Mexique en passant en août au-dessus des prairies montagnardes où il semble qu'elles suivent la floraison des plantes herbacées[27]. Le colibri roux est celui qui effectue la plus longue route migratoire. Il migre de l'Alaska aux régions du nord et du centre du Mexique. Le colibri à gorge rubis est un autre migrateur pouvant traverser d'une seule traite le golfe du Mexique et parcourant ainsi environ 1 000 km sans aucune halte possible.
En général, les mâles arrivent en premier pour établir leur propre territoire. Peu après leur arrivée, les femelles surviennent et déterminent le leur selon le lieu de nidification qui leur semble le plus propice pour élever leur petite famille. Les mâles démontrent beaucoup d'agressivité envers tout autre oiseau-mouche qui voudrait pénétrer dans son territoire, même une femelle. Cette dernière démontre également de l'agressivité mais à un moindre niveau. Elle est trop affairée à la construction du nid et à l'éducation des jeunes. Ces espèces effectuent de courtes haltes où les individus se réservent des territoires dans des endroits qui coïncident avec les territoires de reproduction d'autres espèces. C'est ainsi que l'on peut apercevoir en même temps, sur la côte ouest des États-Unis, le colibri d'Anna, le colibri d’Allen, et le colibri de Costa, en période de reproduction pendant que l'on y retrouve le colibri à queue large et le colibri calliope, durant leur migration.
Certains colibris roux montrent une tendance à migrer d'ouest en est durant l'hiver plutôt que de migrer vers l'Amérique Centrale. Ce phénomène a toujours eu lieu, mais en raison des perspectives alimentaires fournies par les humains, ceux-ci survivent mieux qu'auparavant et transmettent leur route de migration à leur progéniture. Cette espèce se montre assez résistante et si la nourriture et les abris sont suffisants, cette population est capable de tolérer des températures de -20 °C.
Prédateurs
Leurs prédateurs principaux sont les petits félins, les serpents et les autres oiseaux comme les faucons Micrastur et des chevêchettes Glaucidium. Les chats sont même une des causes principales de la disparition des Sephanoides fernandensis au Chili. Leurs prédateurs les plus dangereux sont en fait les espèces qui pillent les nids comme les Corvidae, les Ramphastidae, et quelques micro-chiroptères. Les femelles regagnent leurs nids en volant en zigzags ou en demi-cercles pour éviter de mener un prédateur directement au nid.
Classification, dénomination et histoires
Taxonomie et dénomination
 Aglaeactis castelnaudii par John Gould
Aglaeactis castelnaudii par John GouldLouis Marie Pantaleon Costa réalisa une des premières grandes collections de ces oiseaux au début du XIXe siècle. Jules Bourcier fut un grand spécialiste de ces oiseaux sur la fin de ce siècle. John Gould a publié la célèbre Birds of New Guinea et Monograph of Trochilidae entre 1849-61 et un supplément entre 1880-87.
Le terme Trochilidae a été construit à partir du grec ancien et signifie « petit oiseau ». Selon certaines sources, le terme « colibri » dériverait d'une langue caribe[28]. Dans les classifications traditionnelles, cette famille était classée dans l'ordre des Apodiformes. La classification de Sibley place cette famille dans les Trochiliformes qui est monotypique, mais cette hypothèse est discutée.
Les Nectariniidae qui leur ressemblent par la couleur de leur plumage ou leur alimentation, ne sont que des parents plus lointains (Passeriformes) qui vivent dans l'océan Indien et le Pacifique. Il arrive que certaines de ces espèces soient qualifiées à tort de colibri ou d'oiseau-mouche.
Histoire de l’espèce
Les ailes des oiseaux-mouches sont creuses et fragiles, rendant la fossilisation difficile, l'histoire de cette famille n'est donc connue que grâce aux analyses phylogénétiques. Il semble que les oiseaux-mouches aient un ancêtre commun avec les martinets, il y a environ 35 Ma. Deux fossiles d'espèces très voisines, datés de 29 Ma, ont été mis au jour dans le Bade-Wurtemberg[29] et nommés Eurotrochilus inexpectatus. Ils vivaient dans un climat comparable à celui de l'Amérique du Sud actuellement. Un spécimen de Eurotrochilus bien conservé et trouvé en France révèle que ceux-ci sont morphologiquement très modernes et plus proches des Trochilinae que des Phaethornithinae[30]. Le plus vieux fossile comparable avait été mis au jour en Amérique du Sud et datait de 1 Ma seulement[31].
L'hybridation de l'ADN suggère que l'explosion radiative initiale ait eu lieu, au moins en partie, au cours du Serravallien, entre 12 et 13 Ma, pendant l'émergence des Andes[32]. Ces oiseaux sont passés d'une altitude de 1 500 mètres à 2 000 mètres[33]. On les suppose originaires de l'Amérique du Sud car leur diversité y est la plus importante.
Systématique
On dénombre plus de 330 espèces d'oiseaux-mouches réparties en Amérique tempérée et tropicale.
Dans la classification classique :
─oCaprimulgiformes └─o ├─o │ └─o de nombreuses familles comportant des espèces éteintes et vivantes dont Podargidae, Batrachostomidae, Nyctibiidae └─o Daedalornithes ├─o Aegothelidae └─o ├─o Caprimulgidae └─o Apodiformes ├─o Trochilidae └─o Apodi ├─? Jungornithidae, └─o Laputavis, Aegialornithidae, Hemiprocnidae,
Eocypselus, Apodidae dont Cypseloidinae et ApodinaeListe des espèces
Les Phaethornithinae sont considérés plus primitifs que les Trochilinae par certains auteurs. Les Phaethornithinae ne possèdent pas l'habituel dimorphisme sexuel des oiseaux-mouches, les plumes des mâles et celles des femelles sont iridescentes.
N.B. : l'ordre de cette liste n'est pas aléatoire, il correspond à des liens de parenté entre les différentes espèces (phylogénie). Elle a été établie à partir des listes d'Alan P. Peterson et de la Commission internationale des noms français des oiseaux (Cinfo).
Sous-famille des Phaethornithinae- Ramphodon naevius – Ermite tacheté
- Eutoxeres aquila – Bec-en-faucille aigle
- Eutoxeres condamini – Bec-en-faucille de La Condamine
- Glaucis dohrnii – Ermite de Dohrn
- Glaucis hirsutus – Ermite hirsute
- Glaucis aeneus – Ermite bronzé
- Threnetes ruckeri – Ermite de Rucker
- Threnetes niger – Ermite d'Antonia
- Threnetes leucurus – Ermite à queue blanche
- Anopetia gounellei – Ermite de Gounelle
- Phaethornis yaruqui – Ermite yaruqui
- Phaethornis guy – Ermite vert
- Phaethornis hispidus – Ermite d'Osery
- Phaethornis longirostris – Ermite à longue queue
- Phaethornis superciliosus – Ermite à brins blancs
- Phaethornis malaris – Ermite à long bec
- Phaethornis syrmatophorus – Ermite à ventre fauve
- Phaethornis koepckeae – Ermite de Koepcke
- Phaethornis philippii – Ermite de Filippi
- Phaethornis bourcieri – Ermite de Bourcier
- Phaethornis anthophilus – Ermite anthophile
- Phaethornis eurynome – Ermite eurynome
- Phaethornis pretrei – Ermite de Prêtre
- Phaethornis augusti – Ermite d'Auguste
- Phaethornis subochraceus – Ermite ocré
- Phaethornis squalidus – Ermite terne
- Phaethornis rupurumii – Ermite balafré
- Phaethornis longuemareus – Ermite nain
- Phaethornis idaliae – Ermite d'Idalie
- Phaethornis nattereri – Ermite de Natterer
- Phaethornis ruber – Ermite roussâtre
- Phaethornis stuarti – Ermite de Stuart
- Phaethornis atrimentalis – Ermite à gorge noire
- Phaethornis aethopyga – (?)
- Phaethornis striigularis – Ermite à gorge rayée
- Phaethornis griseogularis – Ermite à gorge grise
Sous-famille des Trochilinae- Androdon aequatorialis – Porte-lance d'Équateur
- Doryfera ludovicae – Porte-lance de Louise
- Doryfera johannae – Porte-lance de Jeanne
- Phaeochroa cuvierii – Colibri de Cuvier
- Campylopterus curvipennis – Campyloptère pampa
- Campylopterus pampa – (?)
- Campylopterus excellens – Campyloptère de Wetmore
- Campylopterus largipennis – Campyloptère à ventre gris
- Campylopterus rufus – Campyloptère roux
- Campylopterus hyperythrus – Campyloptère rougeâtre
- Campylopterus hemileucurus – Campyloptère violet
- Campylopterus ensipennis – Campyloptère à queue blanche
- Campylopterus falcatus – Campyloptère lazulite
- Campylopterus phainopeplus – Campyloptère des Santa Marta
- Campylopterus villaviscensio – Campyloptère du Napo
- Campylopterus duidae – Campyloptère montagnard
- Aphantochroa cirrochloris – Colibri vert et gris
- Eupetomena macroura – Colibri hirondelle
- Florisuga mellivora – Colibri jacobin
- Florisuga fusca – Colibri demi-deuil
- Colibri delphinae – Colibri de Delphine
- Colibri thalassinus – Colibri thalassin
- Colibri coruscans – Colibri anaïs
- Colibri serrirostris – Colibri à ventre blanc
- Anthracothorax viridigula – Mango à cravate verte
- Anthracothorax prevostii – Mango de Prévost
- Anthracothorax nigricollis – Mango à cravate noire
- Anthracothorax veraguensis – Mango de Veraguas
- Anthracothorax dominicus – Mango doré
- Anthracothorax viridis – Mango vert
- Anthracothorax mango – Mango de Jamaïque
- Avocettula recurvirostris – Colibri avocette
- Topaza pella – Colibri topaze
- Topaza pyra – Colibri flamboyant
- Eulampis jugularis – Colibri madère
- Eulampis holosericeus – Colibri falle-vert
- Chrysolampis mosquitus – Colibri rubis-topaze
- Orthorhyncus cristatus – Colibri huppé
- Klais guimeti – Colibri à tête violette
- Stephanoxis lalandi – Colibri de Delalande
- Abeillia abeillei – Colibri d'Abeillé
- Lophornis ornatus – Coquette huppe-col
- Lophornis gouldii – Coquette de Gould
- Lophornis magnificus – Coquette magnifique
- Lophornis brachylophus – Coquette du Guerrero
- Lophornis delattrei – Coquette de Delattre
- Lophornis stictolophus – Coquette pailletée
- Lophornis chalybeus – Coquette chalybée
- Lophornis pavoninus – Coquette paon
- Lophornis helenae – Coquette d'Hélène
- Lophornis adorabilis – Coquette adorable
- Discosura popelairii – Coquette de Popelaire
- Discosura langsdorffi – Coquette de Langsdorff
- Discosura letitiae – Coquette de Letizia
- Discosura conversii – Coquette à queue fine
- Discosura longicaudus – Coquette à raquettes
- Trochilus polytmus – Colibri à tête noire
- Trochilus scitulus – Colibri à bec noir
- Chlorestes notata – Colibri à menton bleu
- Chlorostilbon auriceps – Émeraude couronnée
- Chlorostilbon forficatus – Émeraude de Cozumel
- Chlorostilbon canivetii – Émeraude de Canivet
- Chlorostilbon assimilis – Émeraude du Panama
- Chlorostilbon mellisugus – Émeraude orvert
- Chlorostilbon melanorhynchus – Émeraude des Andes occidentales
- Chlorostilbon gibsoni – Émeraude de Gibson
- Chlorostilbon olivaresi – Émeraude d'Olivares
- Chlorostilbon lucidus – Émeraude splendide
- Chlorostilbon ricordii – Émeraude de Ricord
- Chlorostilbon swainsonii – Émeraude d'Hispaniola
- Chlorostilbon maugaeus – Émeraude de Porto Rico
- Chlorostilbon russatus – Émeraude cuivrée
- Chlorostilbon stenurus – Émeraude à queue étroite
- Chlorostilbon alice – Émeraude alice
- Chlorostilbon poortmani – Émeraude à queue courte
- Panterpe insignis – Colibri insigne
- Elvira chionura – Colibri elvire
- Elvira cupreiceps – Colibri à tête cuivrée
- Eupherusa cyanophrys – Colibri d'Oaxaca
- Eupherusa poliocerca – Colibri du Guerrero
- Eupherusa eximia – Colibri à épaulettes
- Eupherusa nigriventris – Colibri à ventre noir
- Goethalsia bella – Colibri du Pirré
- Goldmania violiceps – Colibri à calotte violette
- Cynanthus sordidus – Colibri sombre
- Cynanthus latirostris – Colibri circé
- Cynanthus doubledayi – Colibri de Doubleday
- Cyanophaia bicolor – Colibri à tête bleue
- Thalurania ridgwayi – Dryade du Mexique
- Thalurania colombica – Dryade couronnée
- Thalurania fannyi – Dryade de Fanny
- Thalurania furcata – Dryade à queue fourchue
- Thalurania watertonii – Dryade de Waterton
- Thalurania glaucopis – Dryade glaucope
- Damophila julie – Colibri de Julie
- Lepidopyga coeruleogularis – Colibri faux-saphir
- Lepidopyga lilliae – Colibri de Lillie
- Lepidopyga goudoti – Colibri de Goudot
- Hylocharis eliciae – Saphir d'Elicia
- Hylocharis sapphirina – Saphir à gorge rousse
- Hylocharis cyanus – Saphir azuré
- Hylocharis chrysura – Saphir à queue d'or
- Hylocharis grayi – Saphir ulysse
- Hylocharis humboldtii – Saphir de Humboldt
- Chrysuronia oenone – Saphir œnone
- Leucochloris albicollis – Colibri à gorge blanche
- Polytmus guainumbi – Colibri guaïnumbi
- Polytmus milleri – Colibri des tépuis
- Polytmus theresiae – Colibri tout-vert
- Leucippus fallax – Colibri trompeur
- Leucippus baeri – Colibri de Tumbes
- Leucippus taczanowskii – Colibri de Taczanowski
- Leucippus chlorocercus – Colibri à queue verte
- Taphrospilus hypostictus – Colibri grivelé
- Amazilia chionogaster – Ariane à ventre blanc
- Amazilia viridicauda – Ariane du Pérou
- Amazilia rutila – Ariane cannelle
- Amazilia yucatanensis – Ariane du Yucatan
- Amazilia tzacatl – Ariane à ventre gris
- Amazilia castaneiventris – Ariane à ventre roux
- Amazilia amazilia – Ariane de Lesson
- Amazilia leucogaster – Ariane vert-doré
- Amazilia versicolor – Ariane versicolore
- Amazilia brevirostris – Ariane à poitrine blanche
- Amazilia franciae – Ariane de Francia
- Amazilia candida – Ariane candide
- Amazilia cyanocephala – Ariane à couronne azur
- Amazilia violiceps – Ariane à couronne violette
- Amazilia viridifrons – Ariane à front vert
- Amazilia wagneri – Ariane de Wagner
- Amazilia fimbriata – Ariane de Linné
- Amazilia lactea – Ariane saphirine
- Amazilia amabilis – Ariane aimable
- Amazilia decora – Ariane charmante
- Amazilia rosenbergi – Ariane de Rosenberg
- Amazilia boucardi – Ariane de Boucard
- Amazilia luciae – Ariane de Lucy
- Amazilia saucerrottei – Ariane de Sophie
- Amazilia cyanifrons – Ariane à front bleu
- Amazilia edward – Ariane d'Edward
- Amazilia cyanura – Ariane à queue bleue
- Amazilia beryllina – Ariane béryl
- Amazilia viridigaster – Ariane à ventre vert
- Amazilia tobaci – Ariane de Félicie
- Microchera albocoronata – Colibri à coiffe blanche
- Anthocephala floriceps – Colibri à tête rose
- Chalybura buffonii – Colibri de Buffon
- Chalybura urochrysia – Colibri à queue bronzée
- Lampornis clemenciae – Colibri à gorge bleue
- Lampornis amethystinus – Colibri à gorge améthyste
- Lampornis viridipallens – Colibri vert-d'eau
- Lampornis sybillae – Colibri de Sybil
- Lampornis hemileucus – Colibri à gorge lilas
- Lampornis calolaemus – Colibri à gorge pourprée
- Lampornis castaneoventris – Colibri à ventre châtain
- Lampornis cinereicauda – Colibri à queue grise
- Basilinna xantusii – Colibri de Xantus
- Basilinna leucotis – Colibri à oreilles blanches
- Lamprolaima rhami – Colibri à gorge grenat
- Adelomyia melanogenys – Colibri moucheté
- Phlogophilus hemileucurus – Colibri à queue mi-blanche
- Phlogophilus harterti – Colibri de Hartert
- Clytolaema rubricauda – Colibri rubis-émeraude
- Heliodoxa xanthogonys – Brillant à couronne verte
- Heliodoxa gularis – Brillant à gorge rose
- Heliodoxa branickii – Brillant de Branicki
- Heliodoxa schreibersii – Brillant à gorge noire
- Heliodoxa aurescens – Brillant à bandeau bleu
- Heliodoxa rubinoides – Brillant rubinoïde
- Heliodoxa jacula – Brillant fer-de-lance
- Heliodoxa imperatrix – Brillant impératrice
- Heliodoxa leadbeateri – Brillant à front violet
- Eugenes fulgens – Colibri de Rivoli
- Hylonympha macrocerca – Colibri à queue en ciseaux
- Sternoclyta cyanopectus – Colibri à poitrine violette
- Urochroa bougueri – Colibri de Bouguer
- Boissonneaua flavescens – Colibri flavescent
- Boissonneaua matthewsii – Colibri de Matthews
- Boissonneaua jardini – Colibri de Jardine
- Aglaeactis cupripennis – Colibri étincelant
- Aglaeactis aliciae – Colibri d'Alice
- Aglaeactis castelnaudii – Colibri de Castelnau
- Aglaeactis pamela – Colibri paméla
- Oreotrochilus chimborazo – Colibri du Chimborazo
- Oreotrochilus estella – Colibri estelle
- Oreotrochilus leucopleurus – Colibri à flancs blancs
- Oreotrochilus melanogaster – Colibri à plastron noir
- Oreotrochilus adela – Colibri adèle
- Lafresnaya lafresnayi – Colibri de Lafresnaye
- Coeligena coeligena – Inca céleste
- Coeligena wilsoni – Inca brun
- Coeligena prunellei – Inca noir
- Coeligena torquata – Inca à collier
- Coeligena phalerata – Inca à queue blanche
- Coeligena bonapartei – Inca de Bonaparte
- Coeligena orina – Inca de Wetmore
- Coeligena helianthea – Inca porphyre
- Coeligena lutetiae – Inca à gemme bleue
- Coeligena violifer – Inca violifère
- Coeligena iris – Inca iris
- Ensifera ensifera – Colibri porte-épée
- Pterophanes cyanopterus – Colibri à ailes saphir
- Patagona gigas – Colibri géant
- Sephanoides sephanoides – Colibri du Chili
- Sephanoides fernandensis – Colibri robinson
- Heliangelus mavors – Héliange mars
- Heliangelus amethysticollis – Héliange de Clarisse
- Heliangelus strophianus – Héliange à queue bleue
- Heliangelus exortis – Héliange tourmaline
- Heliangelus micraster – Héliange menu
- Heliangelus viola – Héliange violette
- Heliangelus zusii – Héliange de Bogota
- Heliangelus regalis – Héliange royal
- Eriocnemis nigrivestis – Érione à robe noire
- Eriocnemis isabellae – Érione d'Isabella
- Eriocnemis vestita – Érione pattue
- Eriocnemis derbyi – Érione de Derby
- Eriocnemis godini – Érione turquoise
- Eriocnemis cupreoventris – Érione à ventre cuivré
- Eriocnemis luciani – Érione de Buquet
- Eriocnemis mosquera – Érione à poitrine d'or
- Eriocnemis glaucopoides – Érione à front bleu
- Eriocnemis mirabilis – Érione multicolore
- Eriocnemis aline – Érione d'Aline
- Haplophaedia aureliae – Érione d'Aurélie
- Haplophaedia assimilis – Érione à pattes rousses
- Haplophaedia lugens – Érione givrée
- Urosticte benjamini – Colibri de Benjamin
- Urosticte ruficrissa – Colibri à sous-caudales rousses
- Ocreatus underwoodii – Haut-de-chausses à palettes
- Lesbia victoriae – Porte-traîne lesbie
- Lesbia nuna – Porte-traîne nouna
- Sappho sparganura – Colibri sapho
- Polyonymus caroli – Colibri de Bourcier
- Ramphomicron dorsale – Colibri à dos noir
- Ramphomicron microrhynchum – Colibri à petit bec
- Oreonympha nobilis – Colibri noble
- Oxypogon guerinii – Colibri casqué
- Metallura tyrianthina – Métallure émeraude
- Metallura iracunda – Métallure dorée
- Metallura williami – Métallure verte
- Metallura baroni – Métallure de Baron
- Metallura odomae – Métallure du Chinguela
- Metallura theresiae – Métallure de Thérèse
- Metallura eupogon – Métallure à gorge feu
- Metallura aeneocauda – Métallure à queue d'airain
- Metallura phoebe – Métallure phébé
- Chalcostigma ruficeps – Métallure à tête rousse
- Chalcostigma olivaceum – Métallure olivâtre
- Chalcostigma stanleyi – Métallure de Stanley
- Chalcostigma heteropogon – Métallure à queue bronzée
- Chalcostigma herrani – Métallure arc-en-ciel
- Opisthoprora euryptera – Colibri avocettin
- Taphrolesbia griseiventris – Colibri comète
- Aglaiocercus kingii – Sylphe à queue d'azur
- Aglaiocercus berlepschi – Sylphe du Venezuela
- Aglaiocercus coelestis – Sylphe à queue violette
- Augastes scutatus – Colibri superbe
- Augastes lumachella – Colibri lumachelle
- Schistes geoffroyi – Colibri de Geoffroy
- Heliothryx barroti – Colibri féerique
- Heliothryx auritus – Colibri oreillard
- Heliactin bilophus – Colibri aux huppes d'or
- Loddigesia mirabilis – Loddigésie admirable
- Heliomaster constantii – Colibri de Constant
- Heliomaster longirostris – Colibri corinne
- Heliomaster squamosus – Colibri médiastin
- Heliomaster furcifer – Colibri d'Angèle
- Rhodopis vesper – Colibri vesper
- Thaumastura cora – Colibri cora
- Tilmatura dupontii – Colibri zémès
- Doricha enicura – Colibri à queue singulière
- Doricha eliza – Colibri d'Eliza
- Calliphlox amethystina – Colibri améthyste
- Calliphlox evelynae – Colibri des Bahamas
- Calliphlox bryantae – Colibri magenta
- Calliphlox mitchellii – Colibri de Mitchell
- Microstilbon burmeisteri – Colibri de Burmeister
- Calothorax lucifer – Colibri lucifer
- Calothorax pulcher – Colibri charmant
- Mellisuga minima – Colibri nain
- Mellisuga helenae – Colibri d'Elena
- Archilochus colubris – Colibri à gorge rubis
- Archilochus alexandri – Colibri à gorge noire
- Calypte anna – Colibri d'Anna
- Calypte costae – Colibri de Costa
- Atthis heloisa – Colibri héloïse
- Atthis ellioti – Colibri d'Elliot
- Myrtis fanny – Colibri de Fanny
- Eulidia yarrellii – Colibri d'Arica
- Myrmia micrura – Colibri à queue courte
- Chaetocercus mulsant – Colibri de Mulsant
- Chaetocercus bombus – Colibri bourdon
- Chaetocercus heliodor – Colibri héliodore
- Chaetocercus astreans – Colibri des Santa Marta
- Chaetocercus berlepschi – Colibri de Berlepsch
- Chaetocercus jourdanii – Colibri de Jourdan
- Selasphorus platycercus – Colibri à queue large
- Selasphorus rufus – Colibri roux
- Selasphorus sasin – Colibri d'Allen
- Selasphorus flammula – Colibri flammule
- Selasphorus ardens – Colibri ardent
- Selasphorus scintilla – Colibri scintillant
- Stellula calliope – Colibri calliope
Les hybrides
Peu d'hybridations ont donné lieu à une étude scientifique. On peut cependant citer celle de Amazilia tzacatl × Amazilia rutila[34], celle de Basilinna leucotis × Selasphorus platycercus[35] ou celle de Selasphorus sasin × Calypte anna[36],[37].
Menaces
Deux espèces sont répertoriées comme éteintes par l'UICN en 2007, Chlorostilbon elegans et Chlorostilbon bracei[38]. Lophornis brachylophus, Lepidopyga lilliae, Amazilia luciae, Amazilia castaneiventris, Aglaeactis aliciae, Coeligena orina, Sephanoides fernandensis, Eriocnemis nigrivestis, Eriocnemis godini, Eriocnemis mirabilis sont en danger critique d'extinction[38].
Oiseaux-mouches et l’homme
Mythes
 L'oiseau-mouche est représenté dans les lignes de Nazca
L'oiseau-mouche est représenté dans les lignes de NazcaLes mythes amérindiens accordent souvent un rôle important aux Trochilidae.
En Mésoamérique, en particulier, le dieu tribal emblématique du peuple aztèque, Huitzilopochtli, était fortement associé aux oiseaux-mouches. De par son nom, d'abord, qui peut se traduire par « oiseau-mouche de gauche » (la gauche symbolisant le sud, traditionnellement associé à la mort), mais aussi parce que, dans la mythologie aztèque, les guerriers morts au combat rejoignaient Huitzilopochtli sous la forme d'un oiseau-mouche butinant des fleurs. De fait, l'oiseau-mouche était un animal sacré chez les Aztèques. On retrouve également dans le nom du souverain aztèque Huitzilihuitl le mot nahuatl désignant l'oiseau-mouche, « huitzil », dont l'origine est une onomatopée imitant le son produit par le vol des oiseaux-mouches.
Chez les Taïnos, l'oiseau-mouche est le symbole du semeur de la vie sur Terre et celui de la renaissance de la nation indienne dans les Caraïbes et la Floride[39].
Les Ohlones racontent que l'oiseau-mouche a apporté le feu aux hommes[40]. Un des géoglyphes de Nazca dépeint un oiseau-mouche.
Culture actuelle
- Trinité-et-Tobago se revendique comme « la terre de l'oiseau-mouche » et un oiseau-mouche figure sur les pièces de monnaie de un cent aussi bien que sur le logo de la compagnie aérienne nationale, Caribbean Airlines.
- Plusieurs chansons populaires en anglais ont eu pour titre « Hummingbird », la traduction pour oiseau-mouche.
- Au Brésil, un bourdonnement d'oiseau-mouche noir est un signe de mort dans la famille, particulièrement s'il a lieu à l'intérieur d'une maison.
Importance économique
Dans le passé, des oiseaux-mouches ont été activement chassés pour leurs plumes qui ont été employées pour faire des bijoux et pour orner des vêtements. Ils ont aussi été capturés pour garnir les zoos et les collections privées. Certains oiseaux-mouches sont toujours vendus sur les marchés comme aphrodisiaques bien qu'aucune preuve d'efficacité dans ce domaine n'ait jamais été démontrée.
Les oiseaux-mouches ont une importance notable sur l'écosystème en étant des pollinisateurs uniques de nombreuses plantes[41]. La présence de ces oiseaux peut attirer les amateurs d'écotourisme.
Engouement du public pour les oiseaux-mouches
 Mangeoire à oiseaux-mouches à San Diego
Mangeoire à oiseaux-mouches à San DiegoLes oiseaux-mouches sont populaires, par leur aspect coloré, leurs chants et le fait qu'ils butinent les fleurs des jardins. Les jardiniers aimant ces oiseaux choisissent les fleurs qu'ils cultivent pour en attirer le plus grand nombre d'espèces.
Ces oiseaux apparaissent d'autant plus fragiles qu'il arrive que les oiseaux-mouches se retrouvent prisonniers à l'intérieur des maisons. Dès lors, leur réflexe est de voler vers le haut, ils ne peuvent donc plus sortir et meurent en moins d'une heure, étant donné leur métabolisme. Par conséquent des mangeoires spécifiques sont disposées pour les nourrir en Amérique du Nord. Ces mangeoires remplacent les jardins de fleurs particulièrement en début et fin de saison. Le nectar synthétique est composé de sucre. Le miel est à éviter car il contient des bactéries nocives pour les oiseaux-mouches[42], les édulcorants trop peu calorifiques sont à proscrire, les colorants sont probablement toxiques[43]. Il est préconisé d'utiliser simplement de l'eau et du sucre[44], de changer le contenu du réservoir régulièrement, en fonction de la chaleur, et de bien le nettoyer à chaque fois pour éviter le développement de bactéries. Des mangeoires à concentration en sucre différentes peuvent permettre aux oiseaux-mouches de différentes tailles de créer une hiérarchie qui permettra aux plus petits d'accéder aux mangeoires confisquées par les plus gros.
Voir aussi
Bibliographie
- (en) Dennis John V. et Mathew Tekulsky, How to Attract Hummingbirds & Butterflies, San Ramon, Ortho Books, 1991
- Pomarède Maurice, Colibris, les cupidons des Amériques, Sciences & nature, 1991, chap. 14
- Stokes Donald W. et Lillian Q. Stokes, The Hummingbird Book, Toronto, Little, Brown and Company, 1989
- (en)Why are hummingbirds so small, so beautiful and so different from other birds?, une légende Maya sur l'origine des oiseaux-mouches
- (en) Erich D. Jarvis, Sidarta Ribeiro, Maria Luisa da Silva, Dora Ventura, Jacques Vielliardk & Claudio V. Mello, « Behaviourally driven gene expression reveals song nuclei in hummingbird brain », dans Nature, vol. 406, août 2000 [texte intégral]
Source
- (fr)Une partie de ce texte est reprise de Rober Morin, « Le monde fascinant des oiseaux mouches » sur coq.qc.ca. Suivant la licence spécifiée, le contenu du texte peut être reproduit à condition d'en mentionner la source.
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Hummingbird » (voir la liste des auteurs)
Références externes
 Femelle de Lampornis castaneoventris.
Femelle de Lampornis castaneoventris.- Référence Alan P. Peterson : Trochilidae dans Apodiformes (en)
- Référence Catalogue of Life : Trochilidae (en)
- Référence The Paleobiology database : Trochilidae Vigors 1825 (en)
- Référence ITIS : Trochilidae (fr) ( (en))
- Référence Animal Diversity Web : Trochilidae (en)
- Référence NCBI : Trochilidae (en)
Liens externes
- Référence Oiseaux.net : Trochilidae (fr)
- Référence UICN : taxon Trochilidae (en)
- Référence CITES : famille Trochilidae (sur le site de l’UNEP-WCMC) (fr+en)
- John A. Gerwin, and Robert M. Zink, « Phylogenic patterns in the Trochilidae », dans The Auk, Museum of Natural Science and Department of Zoology and Physiology, Louisiana State University, vol. 115, no 1, 1998, p. 105-118 [texte intégral]
- Les photos d'oiseaux-mouches
- Video (.wmv) (.mp4 - iPod) Hummingbird hovers and feeds. Licence : Creative Commons Attribution-ShareAlike. Disponible sur blip.tv
- Video (.wmv) Hummingbird shaping a nest. Licence Creative Commons Attribution-ShareAlike. Disponible sur blip.tv
- Video (.wmv) Hummingbird adding spider's web to nest. Licence Creative Commons Attribution-ShareAlike. Disponible sur blip.tv
- Video (.wmv) Hummingbird adding downy seed to nest. License Creative Commons Attribution-ShareAlike. Disponible sur blip.tv
- (en)Erol Cagatay, George Chi, et Jenna Harsch, « Study of Hummingbird Aerodynamics in Relation to Micro Air Vehicles », Université du Texas, 1998, sur le vol des oiseaux-mouches
- (en)Hummingbird Studies Raise Questions About Birdsong Evolution, Science Daily
- (en)Trochilidae, BIRD FAMILIES OF THE WORLD sur montereybay.com
Notes
- Groupe d'étude et de protection des oiseaux en Guyane (2003) Portraits d'oiseaux guyanais. Ibis Rouge Editions, Guyane, Guadeloupe, Martinique, Paris, Réunion, 479 p.
- locomotion, Colibri sur museevirtuel.ca
- (fr)Le plumage et cycles sur oiseau.net
- (Alaine Camfield, Kari Kirschbaum sur ADW, 2007)
- (en)Birds: Hummingbird, Zoo de San Diego
- La langue, Colibri sur museevirtuel.ca
- (en) Fenster C. B, « Selection on floral morphology by hummingbirds », dans Biotropica, vol. 23, no 1, 1991, p. 98-101 (ISSN 0006-3606) [résumé, texte intégral]
- (en) F. Gary Stiles, « Geographical Aspects of Bird-Flower Coevolution, with Particular Reference to Central America », dans Annals of the Missouri Botanical Garden, vol. 68, no 2, 1981, p. 323-351 [résumé]
- (en) Paton,D.C. et B.G. Collins, « Bills and tongues of nectar-feeding birds:A review of morphology, function, and performance, with intercontinental comparisons », dans Australian journal of ecology, vol. 14, no 4, 1989, p. 473-506 (ISSN 0307-692X) [résumé]
- Omara-Otunnu, Elizabeth. Hummingbird's Beaks Bend To Catch Insects. University of Connecticut Advance (2004-07-19).
- (en) EBERHARD J. R. ; EWALD P. W., « Food availability, intrusion pressure and territory size: an experimental study of Anna's hummingbirds (Calypte anna) », dans Behavioral ecology and sociobiology, vol. 34, no 1, 1994, p. 11-18 (ISSN 0340-5443) [résumé]
- (en) Erich D. Jarvis, Sidarta Ribeiro, Maria Luisa da Silva, Dora Ventura, Jacques Vielliardk & Claudio V. Mello, « Behaviourally driven gene expression reveals song nuclei in hummingbird brain », dans Nature, vol. 406, août 2000 [texte intégral]
- (en) DEVOTO Mariano; MONTALDO Norberto H.; MEDAN Diego, « Mixed hummingbird : Long-proboscid-fly pollination in 'ornithophilous' Embothrium coccineum (Proteaceae) along a rainfall gradient in Patagonia, Argentina », dans Austral ecology, vol. 31, no 4, 2006, p. 512-519 (ISSN 1442-9985) [résumé]
- (en)Gary Ritchison, « Avian Circulatory System », Department of Biological Sciences Eastern Kentucky University
- Skutch, Alexander F. & Singer, Arthur B. (1973): The Life of the Hummingbird. Crown Publishers, New York. ISBN 0-517-50572-X
- (en)Erol Cagatay, George Chi, et Jenna Harsch, « Background Theory » sur Study of Hummingbird Aerodynamics in Relation to Micro Air Vehicles, Université du Texas, 1998
- Système respiratoire, colibris sur museevirtuel.ca
- (en) Preest, M. R. and Beuchat, C. A., « Ammonia excretion by hummingbirds », dans Nature, vol. 386, 1997, p. 561-562
- (en) Todd J. McWhorter, Donald R. Powers, and Carlos Martínez del Rio, « Are Hummingbirds Facultatively Ammonotelic? Nitrogen Excretion and Requirements as a Function of Body Size », dans Physiological and Biochemical Zoology, Université de Chicago, vol. 76, 2003, p. 731–743 [résumé]
- Rayner, J.M.V. 1995. Dynamics of vortex wakes of flying and swimming vertebrates. J. Exp. Biol. 49:131–155.
- (en) Robert S. Ridgely and Paul G. Greenfield, The Birds of Ecuador, Cornell University Press, coll. « Field Guide », 2001
- (en) « Aerodynamics of the hovering hummingbird », dans Nature, vol. 435, 23 juin 2005, p. 1094-1097 [résumé]
- (en) KODRIC-BROWN A.; BROWN J. H. ; BYERS G. S. ; GORI D. F., « Organization of a tropical island community of humming-birds and flowers », dans Ecology, Ecological Society of America, Washington, vol. 65, no 5, 1984, p. 1358-1368 (ISSN 0012-9658) [résumé]
- (en) LARA Carlos; ORNELAS Juan Francisco, « Effects of nectar theft by flower mites on hummingbird behavior and the reproductive success of their host plant, Moussonia deppeana (Gesneriaceae) », dans Oikos, vol. 96, no 3, 2002, p. 470-480 (2 p.) (ISSN 0030-1299) [résumé]
- (en) LARA C.; ORNELAS J. F., « Nectar 'theft' by hummingbird flower mites and its consequences for seed set in Moussonia deppeana », dans Functional ecology, vol. 15, no 1, 2001, p. 78-84 (1 p.1/4) (ISSN 0269-8463) [résumé]
- (en) VENZAL J. M.; FELIX M. L.; OLMOS A.; MANGOLD A. J.; GUGLIELMONE A. A., « A collection of ticks (Ixodidae) from wild birds in Uruguay », dans Experimental & applied acarology, vol. 36, no 4, 2005, p. 325-331 (ISSN 0168-8162) [résumé]
- (en) Curry-Lindahl K., Les Oiseaux migrateurs à travers mer et terre, Neuchâtel, Paris, Delachaux et Niestlé, 1980, 241 p p.
- Définitions lexicographiques et étymologiques de « Colibri » du CNRTL.
- Oldest hummingbird fossil found, 2004
- (en) Antoine Louchart, Nicolas Tourment, Julie Carrier, Thierry Roux et Cécile Mourer-Chauviré, « Hummingbird with modern feathering: an exceptionally well-preserved Oligocene fossil from southern France », dans Naturwissenschaften, vol. 95, février 2008, p. 171-175 (ISSN 0028-1042) [résumé]
- (en) Erik Stokstad, « Old World Fossil Record of Modern-Type Hummingbirds », dans Science magazine, vol. 304, no 5672, 7 mai 2004 [résumé]
- (en)[PDF]Gerwin, John A. & Zink, Robert M., « Phylogenetic patterns in the Trochilidae », dans The Auk, vol. 115, no 1, 1998, p. 105-118 [résumé]
- (en) François Vuilleumier, « Pleistocene Speciation in Birds living in the High Andes », dans Nature, vol. 223, 13 septembre 1969, p. 1179 - 1180 [résumé]
- (en) GRAVES Gary R., « Diagnoses of hybrid hummingbirds (Aves: Trochilidae). 12: Amazilia bangsi Ridgway, 1910, is an intrageneric hybrid, Amazilia tzacatl × Amazilia rutila », dans Proceedings of the Biological Society of Washington, vol. 116, no 4, 2003, p. 847-852 (ISSN 0006-324X) [résumé]
- (en) GRAVES Gary R., « Diagnoses of hybrid hummingbirds (Aves: Trochilidae). 15. A new intergeneric hybrid (Hylocharis leucotis × Selasphorus platycercus) from the Huachuca Mountains, southeastern Arizona », dans Proceedings of the Biological Society of Washington, vol. 120, no 1, ???, p. 99–105 [résumé]
- [PDF](en) Robert Ridgway, « Hybridism and Generic Characters in the Trochilidae. », dans The Auk, 4e série, no 26, 1909, p. 440-442. [texte intégral]
- [PDF](en) Walter P Taylor, « An instance of hybridization in hummingbirds, with remarks on the weight of generic characters in the Trochilidae », dans The Auk, 3e série, no 26, 1909, p. 291-293 [texte intégral]
- (UICN, 2007)
- (en)Hummingbirds in Native American Tradition
- (en) Linda Yamane, When the World Ended, How Hummingbird Got Fire, How People Were Made: Rumsien Ohlone Stories, Oyate, janvier 1995 (ISBN 978-0962517518)
- Temeles E, Linhart Y, Masonjones M & Masonjones H, « The Role of Flower Width in Hummingbird Bill Length–Flower Length Relationships », dans Biotropica, vol. 34, no 1, 2002, p. 68-80 [texte intégral]
- (en)Feeders and Feeding Hummingbirds
- Please Don't Use Red Dye
- Shackelford, Clifford Eugene; Lindsay, Madge M. & Klym, C. Mark, Hummingbirds of Texas with their New Mexico and Arizona ranges, Texas A&M University Press, College Station, 2005 (ISBN 1-58544-433-2)

La version du 25 novembre 2007 de cet article a été reconnue comme « bon article », c'est-à-dire qu'elle répond à des critères de qualité concernant le style, la clarté, la pertinence, la citation des sources et l'illustration. Catégories :- Trochilidae
- Faune endémique du continent américain


















Wikimedia Foundation. 2010.