- Glycolyse
-
La glycolyse (γλῠκὖς glykýs « sucré » et λύσις lýsis « dissolution ») ou voie d'Embden-Meyerhof-Parnas est une voie métabolique d'assimilation du glucose et de production d'énergie. Elle se déroule dans le cytoplasme (ou cytosol) de la cellule. Comme son nom l'indique elle nécessite du glucose et a pour produit du pyruvate. Ce dernier peut soit entrer dans le cycle de Krebs, qui se déroule dans la mitochondrie des eucaryotes ou le cytoplasme des bactéries en aérobiose, soit être métabolisé par fermentation en anaérobiose, pour produire par exemple du lactate ou de l'éthanol.
Principe général
La glycolyse est un mécanisme de régénération d'ATP qui ne nécessite pas d'oxygène. Au cours de ce processus, on assiste à :
- des réactions d'oxydo-réduction au cours desquelles un accepteur d'électrons (coenzyme NAD+) est réduit :
- des synthèses d'ATP par phosphorylation d'ADP (formation de quatre molécules d'ATP, mais consommation de deux molécules d'ATP, soit au total formation nette de deux molécules d'ATP) :
Le symbole Pi représente ici le phosphate inorganique HPO42-, ou hydrogénophosphate[1].
La glycolyse se traduisant par la réduction de coenzymes, elle s'accompagne donc de l'oxydation de molécules organiques. On peut dire qu'elle correspond à l'oxydation du glucose en pyruvate :
- glucose + 2 NAD+ → 2 CH3-CO-COO- + 2 (NADH + H+ ),
couplée à :
soit au total
- * Le pyruvate CH3-CO-COO- désigne en toute rigueur la base conjuguée de l'acide pyruvique CH3-CO-COOH.
Étapes de la glycolyse
Activation des hexoses par phosphorylations successives
Phosphorylation du glucose en glucose-6-phosphate
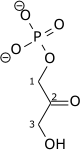
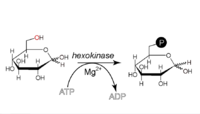 Synthèse du α-D-Glu-6-P.
Synthèse du α-D-Glu-6-P.
Cette réaction nécessite un cation Mg2+ comme cofacteur et consomme une molécule d'ATP pour phosphoryler chaque molécule de glucose. Elle contribue à maintenir la concentration en glucose relativement basse dans le cytoplasme afin de faciliter l'entrée de molécules de glucose supplémentaires. De surcroît, le glucose-6-phosphate ne peut plus quitter la cellule, car la membrane plasmique n'a pas de transporteur pour le Glc-6-P.
- Cette réaction est irréversible. Elle est catalysée par une kinase, soit une hexokinase, non spécifique du glucose qui, chez les mammifères, se trouve le plus souvent dans le muscle, soit une glucokinase, spécifique du glucose. Ces deux enzymes ont des constantes de Michaelis (KM) différentes avec pour valeurs respectives 0,1 mM et 10 mM en sachant que la KM est inversement proportionnelle a l'affinité de l'enzyme pour ses substrats. Ces deux enzymes sont Mg2+ dépendantes. Chez l'homme, la glucokinase est localisée dans le foie et dans les cellules pancréatiques. En effet, cette dernière est parfaitement adaptée à la fonction de stockage du foie (elle fonctionne principalement lors d'afflux de glucose importants, après un repas par exemple, et contribue ainsi à la régulation de la glycémie). Un dysfonctionnement de cette enzyme est donc responsable de certains types de diabète (diabètes MODY qui, pour 50 % des cas, sont dus à une mutation de la glucokinase).
- La phosphorylation du glucose n'est pas spécifique de la glycolyse. Cette étape sert également de point de départ dans la voie des pentoses phosphates ou pour la glycogènogenèse.
Remarque : toutes les reactions qui ont une variation d'enthalpie libre élevée sont irréversibles, et comme cette phosphorylation est énergétiquement très favorisée, la réaction est irréversible. C'est pourquoi ces enzymes sont très régulées afin d'éviter l'emballement du système, à l'instar des deux autres étapes irréversibles de la glycolyse. (Phosphofructokinase-1, Pyruvate kinase). L'hexokinase est notamment inhibée par son propre produit, le glucose-6-phosphate (rétrocontrôle négatif), et son expression génique est induite par l'insuline. La glucokinase n'est quant à elle pas inhibée par le glucose-6-phosphate, mais son expression génique est induite par l'insuline.
Isomérisation du glucose-6-phosphate en fructose-6-phosphate
Le α-D-glucose-6-phosphate produit au cours de la glycolyse est isomérisé en β-D-fructose-6-phosphate par la glucose-6-phosphate isomérase. Cette réaction est réversible, et demeure orientée vers la droite en raison de la concentration en Fru-6-P, maintenue assez faible en raison de sa consommation immédiate par l'étape suivante de la glycolyse.
Phosphorylation du fructose-6-phosphate en fructose-1,6-diphosphate
-
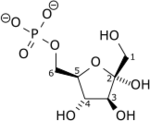
+ ATP → ADP + H+ + 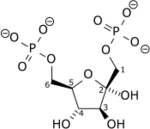
Fru-6-P Fru-1,6-DP Phosphofructokinase-1 – EC
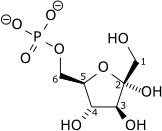
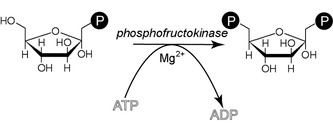 Formation du β-D-fructose-1,6-diphosphate.
Formation du β-D-fructose-1,6-diphosphate.
Le β-D-fructose-6-phosphate (Fru-6-P) produit au cours de la glycolyse est phosphorylé en β-D-fructose-1,6-diphosphate (Fru-1,6-DP) par la phosphofructokinase-1 à partir d'une molécule d'ATP, convertie en ADP. Cette consommation d'énergie rend cette étape irréversible, et constitue un point de régulation majeur de la vitesse de la glycolyse. Un cation Mg2+ intervient comme cofacteur.
Il existe, essentiellement chez des organismes autres que les animaux, des enzymes différentes capables de phosphoryler le Fru-6-P à partir de pyrophosphate inorganique au lieu d'ATP. C'est le cas de la diphosphate fructose-6-phosphate 1-phosphotransférase (PFP), qu'on trouve chez de nombreux végétaux, certaines bactéries, des archées et des protistes. De rares archées possèdent une variante de la phosphofructokinase utilisant, cette fois, de l'ADP et non de l'ATP.
Cette réaction, catalysée par une phosphofructokinase (PFK) est irréversible et Mg2+ dépendante. Cette enzyme catalyse la première étape qui soit spécifique de la glycolyse. Elle est très fortement régulée de manière allostérique par l'ATPlibre (l'ATPlibre est la forme de l'ATP non complexé au magnésium), qui est le produit final "utile" de la glycolyse. Plus la concentration en ATPlibre est importante, plus cette réaction est lente et, inversement, plus la concentration en ATPlibre est faible, plus l'enzyme est active. Il s'agit d'un système cybernétique d'autocontrôle de la glycolyse. Plusieurs modéles mathématiques de la glycolyse ont été mis au point et montrent que cette étape est la plus importante de celles qui contrôlent le flux de la glycolyse.
L'inhibition par l'ATP est réversible par l'AMP, ce qui permet de garder un rapport ATP/AMP constant.
Mais elle est surtout régulée par le fructose-2,6-diphosphate (Fru-2,6-DP) : en effet, la production de Fru-2,6-DP à partir du Fru-6-P a pour seule fonction de mettre en évidence une saturation de la voie en Fru-6-P (« trop plein »), car le Fru-2,6-DP n'a pas de devenir métabolique. Par allostérie, le Fru-2,6-DP active donc la phosphofructokinase-1 afin de stimuler la consommation de Fru-6-P et ainsi empêcher sa propre formation.
Clivage du fructose-1,6-diphosphate en deux molécules de glycéraldéhyde-3-phosphate
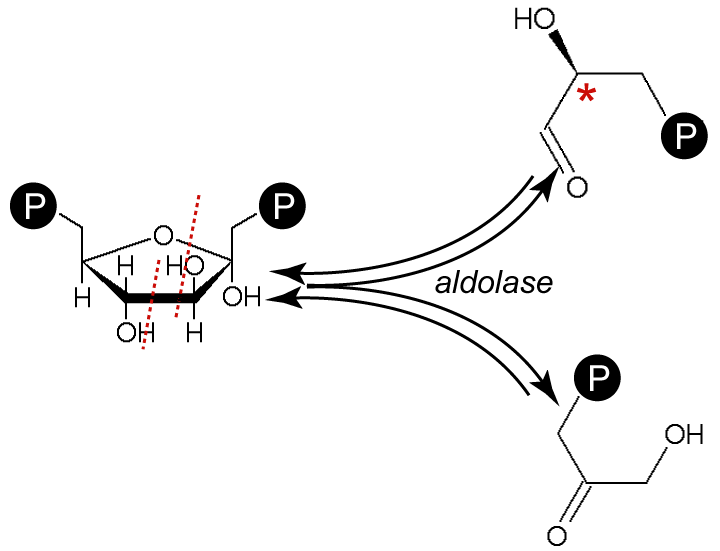
Clivage du fructose-1,6-diphosphate en glycéraldéhyde-3-phosphate et dihydroxyacétone phosphate
Le β-D-fructose-1,6-diphosphate est clivé par une lyase, la fructose-diphosphate aldolase, en D-glycéraldéhyde-3-phosphate (G3P) et dihydroxyacétone phosphate (DHAP).
Il existe deux classes d'aldolases susceptibles de cliver le Fru-1,6-DP : la classe I chez les animaux et les végétaux, et la classe II chez les mycètes et les bactéries ; ces deux classes d'enzymes utilisent des mécanismes différents pour cliver ce cétose.
Isomérisation de la dihydroxyacétone phosphate en glycéraldéhyde-3-phosphate
La dihydroxyacétone phosphate est isomérisée en D-glycéraldéhyde-3-phosphate par la triose-phosphate isomérase. Ainsi, chaque molécule de β-D-fructose-1,6-diphosphate donne en fin de compte deux molécules de D-glycéraldéhyde-3-phosphate.
Cette réaction est peu favorisée, elle a lieu à raison de 5% dans le sens "dihydroxyacétonephosphate → glycéraldéhyde-3-phosphate" et à 95% dans l'autre sens.[réf. nécessaire]
Récupération de l'énergie investie dans les phosphorylations
Phosphorylation du glycéraldéhyde-3-phosphate en 1,3-diphosphoglycérate
Le D-glycéraldéhyde-3-phosphate est phosphorylé en 1,3-diphospho-D-glycérate (1,3-DPG) par la glycéraldéhyde-3-phosphate déshydrogénase avec réduction concomitante d'une molécule de NAD+ en NADH + H+ ; c'est la seule étape de la glycolyse où est formé du pouvoir réducteur, sous forme de NADH + H+. Cette réaction est équilibrée du point de vue de la charge électrique et du nombre d'atomes d'hydrogène par le fait que le phosphate inorganique (Pi) existe, dans le milieu cytoplasmique, sous forme d'ion hydrogénophosphate HPO42-.
Cette réaction d'oxydoréduction, réversible et catalysée par une oxydo-réductase, conduit à la formation d'une liaison acylthioester à haut potentiel de transfert. Cette étape constitue le début de la seconde partie de la glycolyse. L'énergie contenue dans les liaisons à haut potentiel de transfert va être utilisé pour la synthèse de l'ATP. Les coenzymes sont réduites (gain d'électrons).
Conversion du 1,3-diphosphoglycérate en 3-phosphoglycérate avec récupération d'ATP
Le groupe phosphate à haut potentiel de transfert du 1,3-diphospho-D-glycérate (1,3-DPG) permet de phosphoryler une molécule d'ADP en ATP pour former le 3-phospho-D-glycérate (3PG) sous l'action de la phosphoglycérate kinase ; c'est la première étape de la glycolyse où de l'énergie est récupérée sous forme réutilisable, emmagasinée dans l'ATP.
Isomérisation du 3-phosphoglycérate en 2-phosphoglycérate
Le 3-phospho-D-glycérate est isomérisé en 2-phospho-D-glycérate (2PG) par la phosphoglycérate mutase.
Conversion du 2-phosphoglycérate en phosphoénolpyruvate
Le 2-phospho-D-glycérate (2PG) est déshydraté par une lyase, l’énolase (ou phosphopyruvate hydratase), pour former le phosphoénolpyruvate (PEP). Un cation Mg2+ est requis comme « catalyseur » de la réaction de déshydratation, tandis qu'un second Mg2+ intervient avec un rôle « conformationnel » en coordination avec le groupe carboxyle du 2-phospho-D-glycérate.
Conversion du phosphoénolpyruvate en pyruvate avec récupération d'ATP
Le groupe phosphate à haut potentiel de transfert (ΔG°' = -61,9 kJ⋅mol-1) du phosphoénolpyruvate permet la phosphorylation d'une molécule d'ADP en ATP par la pyruvate kinase. Un cation Mg2+ est nécessaire à cette réaction comme cofacteur.
Bilan de la glycolyse

On utilise :
- 1 mole de glucose
- 2 moles de coenzymes oxydés
- 2 moles d'ADP
- 2 moles de Pi (phosphate inorganique)
Pour produire :
- 2 moles de pyruvate
- 2 moles de coenzymes réduits
- 2 moles d'ATP
- 2 moles d'eau
on gagne également 4 protons(H+) : 2 lorsque 2NAD+ DONNE 2NADH + 2H+, 1 lorsque glucose devient glucose-6-phosphate et 1 lorsque fructose-6-phosphate devient fructose-1,6-diphosphate.
On a finalement produit 2 moles d'ATP pour lyser 1 mole de glucose. Ce bilan est faible.
Régulation de la glycolyse
La glycolyse est principalement régulée au niveau de 3 enzymes clés qui sont la PFK-1, la pyruvate kinase et l'hexokinase
Régulation de la PFK-1
La PFK-1 est régulée de façon allostérique:
- L'ATP et le citrate agissent comme des inhibiteurs
- L'AMP et le F 2,6 di-P agissent comme des activateurs.
La concentration en F 2,6 di-P est donc primordiale sur la glycolyse. Elle est régulée par la phosphofructokinase-2 dont l'activité sera différente selon son état de phosphorylation:
- Par l'action du glucagon (hormone hyperglycémiante), elle sera déphosphorylée et catalysera la réaction F 2,6 di-P + H2O → F6P + Pi. Ainsi la concentration de F 2,6 di-P diminuera et la glycolyse sera ralentie.
- Par l'action de l'insuline (hormone hypoglycémiante) elle sera phosphorylée et catalysera la réaction F6P + ATP → F 2,6 di-P + ADP. Ainsi la concentration de F 2,6 di-P augmentera et la glycolyse sera accélérée.
Régulation de la pyruvate kinase
La pyruvate kinase est régulée allostériquement et ceci de façon ubiquitaire :
- L'AMP et le fructose-1,6-diphosphate sont des activateurs
- L'ATP, l'acétyl-CoA et l'alanine sont des inhibiteurs.
Au niveau du foie, elle est également régulée de façon covalente (par l'action d'hormones)
- Le glucagon va phosphoryler cette enzyme pour l'inhiber
- L'insuline va réaliser l'action inverse pour l'activer.
Régulation au niveau de l'hexokinase
L'activité de cette enzyme est inhibée par le produit de la réaction, le glucose 6-phosphate. Si celui-ci s'accumule, sa production va très vite diminuer et s'équilibrer avec sa consommation. Ce processus évite l'accumulation de produits intermédiaires.
Réoxydation des coenzymes
Il est important de comprendre que la glycolyse cesse lorsque les coenzymes ne sont pas réoxydées sous la forme NAD+. Sans ces coenzymes, l'étape catalysée par l'enzyme D-glycéraldéhyde-3-phosphate déshydrogénase ne peut se produire, provoquant l'arrêt de la glycolyse. Il est donc crucial de régénérer ces coenzymes.
Il existe deux voies métaboliques principales pour cela : l'une se fait par phosphorylation au niveau du substrat, et est appelée fermentation. Il en existe de plusieurs sortes : fermentation lactique (qui se produit dans le muscle non oxygéné), fermentation butyrique, alcoolique… L'autre voie de réoxydation des coenzymes se fait par phosphorylation oxydative (par exemple en utilisant l'oxygène comme accepteur d'électron final) et est appelée respiration, certains parlent de respiration cellulaire pour la différencier de la ventilation pulmonaire, bien que les contextes d'utilisation ne prêtent pas à confusion. Elle a lieu au niveau de la chaîne respiratoire des mitochondries (phosphorylation oxydative) chez les eucaryotes et dans le cytoplasme des bactéries. Le bilan énergétique de la glycolyse suivie de la respiration (36 ATP) est environ 20 fois plus élevé que celui de la glycolyse suivie de la fermentation (2 ATP pour la fermentation lactique).
Notes et références
- Biologie moléculaire de la cellule Par Harvey Lodish, Arnold Berk, Paul Matsudaira, James Darnell, Chris A. Kaiser, Pierre L. Masson - page 301.
Voir aussi
Articles connexes
Autres voies de dégradation du glucose :
- la voie des pentoses phosphates
- la voie d'Entner-Doudoroff
- Pyruvate
- la voie inverse de la glycolyse, la néoglucogenèse
Liens externes
- La logique chimique de la glycolyse (anglais)
- Une animation simplifiée du processus de glycolyse (français)












Wikimedia Foundation. 2010.