- Sahelanthropus tchadensis
-
Sahelanthropus tchadensis
Sahelanthropus
tchadensis
Moulage du crâne holotype non-reconstruit
de Sahelanthropus tchadensis TM 266-01-060-1,
surnommé Toumaï, en vue facio-latérale.Classification classique Règne Animalia Embranchement Chordata Sous-embr. Vertebrata Classe Mammalia Ordre Primates Famille Hominidae Sous-famille Homininae Tribu Hominini Genre Sahelanthropus
Brunet et al., 2002Nom binominal Sahelanthropus tchadensis
Brunet et al., 2002Lieu de découverte 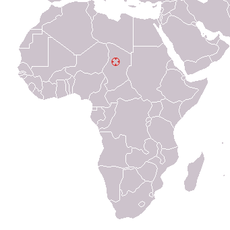
Djourab, Tchad
 Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsParcourez la biologie sur Wikipédia : 





Sahelanthropus tchadensis est le nom donné à un ensemble de fossiles appartenant à une espèce éteinte de primate, considérée par une partie seulement de la communauté scientifique comme un hominine[1] bipède. Le premier spécimen fossile, surnommé « Toumaï », a été découvert au Tchad en juillet 2001 par l'équipe de Michel Brunet dans des niveaux géologiques dont l'âge est compris entre 6,8 et 7,2 millions d'années, selon les résultats obtenus en 2008 par une méthode de datation absolue. Toutefois la méthode s'apparente dans ce cas à un raisonnement de type circulaire d'autant que selon toute vraisemblance le crâne faisait partie d'une tombe récente que le vent venait de dégager.
Il s'agirait pour certains auteurs de l'une des premières espèces de la lignée humaine, probablement très proche de la divergence chimpanzés-hominines. La date de cette divergence est estimée entre -6 et -5 millions d'années par la majorité des études moléculaires, ce qui est donc en désaccord avec l'hypothèse d'un hominine aussi ancien.
En tout état de cause, cette découverte a contribué à la remise en question de l'East Side Story, une théorie popularisée par Yves Coppens selon laquelle l'émergence de la lignée humaine a eu lieu à l'est de la vallée du Rift africain, en relation avec des changements climatiques.
Étymologie
La dénomination générique Sahelanthropus est formée à partir de Sahel, région de la découverte, et de anthropos qui veut dire homme en grec ; elle signifie donc littéralement « homme du Sahel ». La dénomination spécifique tchadensis fait référence au Tchad, pays dans lequel fut trouvé cet hominine.[2]
Toumaï est le surnom donné au premier spécimen découvert, un crâne sub-complet. Il a fréquemment valeur de dénomination vernaculaire pour l’espèce Sahelanthropus tchadensis. Toumaï signifie « espoir de vie » en langue goran[2],[3]. Ce nom a été choisi par le président de la République du Tchad. Il désigne dans cette langue des enfants nés juste avant la saison sèche et qui ont, de ce fait, des chances de survie assez limitées. Mais Toumaï, nom plutôt rare, était aussi celui d'un compagnon de route de l'actuel chef de l'État tchadien qui perdit la vie au cours d'une bataille[4].
Conditions de la découverte
 Michel Brunet
Michel Brunet Article détaillé : Recherche paléoanthropologique au Tchad.
Article détaillé : Recherche paléoanthropologique au Tchad.La découverte de Sahelanthropus tchadensis est le fruit d’une recherche de plusieurs années menée par Michel Brunet, paléoanthropologue, professeur à l’université de Poitiers, nommé fin 2007 professeur au Collège de France[5]. Dès les années 1980, il explore l’Afrique de l'Ouest et notamment le Cameroun à la recherche de couches sédimentaires âgées de 6,5 à 7 millions d’années, afin de tester scientifiquement la théorie de l’« East Side Story ». Cette théorie, popularisée par Yves Coppens en 1982, met en relation l’apparition des premiers hominines en Afrique de l'Est avec les changements climatiques et environnementaux liés à la formation du Grand rift.
Le Centre national d’appui à la recherche (CNAR) du ministère de l’Enseignement supérieur de la République du Tchad l’invite pour une conférence en 1992 et lui propose de participer à des missions de terrain. Sa première mission au Tchad aura lieu en janvier 1994.
Le [21 janvier 1995][6], lors de sa seconde mission, Mamelbaye Tomalta, un chauffeur de la Direction des Mines, découvre dans le secteur de Koro-Toro, en plein désert du Djourab, au nord du Tchad, un reste d'hominine. Il s'agit d'un fragment de mandibule ayant appartenu à un individu qui sera surnommé Abel et qui conduira à la définition de l'espèce Australopithecus bahrelghazali[7]. La Mission paléoanthropologique franco-tchadienne (MPFT), une collaboration scientifique entre l'université de Poitiers, le CNAR et l'université de Ndjamena, naît de cette découverte. Des prospections et fouilles systématiques se développent et permettent la découverte en 1997 du secteur fossilifère de Toros-Menalla, plus de 150 km à l'ouest de Koro-Toro. Sur le 266ème site découvert dans ce secteur, une équipe de quatre hommes en service au CNAR – composée d'Alain Beauvilain, géographe docteur d’État, Fanoné Gongdibé, ingénieur détaché du ministère des Mines, Ahounta Djimdoumalbaye, licencié en sciences naturelles, vacataire au CNAR, et Mahamat Adoum, non diplômé, employé au CNAR – met au jour le [19 juillet 2001][8] le crâne de Sahelanthropus tchadensis qui sera surnommé Toumaï[2].
Fossiles découverts
À ce jour, l’ensemble des fossiles pour l’espèce (hypodigme) se limite à neuf restes crâniens et un reste post-crânien correspondant au minimum à six individus. Ces restes proviennent de trois sites, TM 266, TM 247 et TM 292, situés à quelques kilomètres les uns des autres dans le secteur de Toros-Menalla. Le reste post-crânien est un fémur gauche d'hominidé, parfaitement identifiable, trouvé par Beauvilain le 19 juillet 2001 à côté du crâne. Des nomades, auteurs d'une vraisemblable inhumation du crâne, avaient disposé ce fémur en position d'humérus.
Tous ces fossiles étaient contenus dans la même couche sédimentaire : l’unité à anthracothères (Anthracotheriid Unit : AU)[9].
Ces fossiles, découverts entre juillet 2001 et mars 2002, sont[2],[10] :
- Un crâne sub-complet, holotype de l’espèce référencé sous le code TM 266-01-060-1, qui présente une déformation (compression supéro-inférieure et écrasement du côté gauche associés à des fissures et déformations plastiques). Il s’agit du premier reste découvert le 19 juillet 2001 par Ahounta Djimdoumalbaye. Ce crâne est surnommé Toumaï.
- quatre mandibules partielles ou fragmentaires dont deux publiées en 2002 dans la première publication de l’espèce (TM 266-01-060-2 : fragment de symphyse avec alvéoles de canine et d’incisive, TM 266-02-154-1 : fragment droit avec une P3 fragmentaire, et la suite P4-M3) et deux publiées en 2005 (TM 292-02-01 : mandibule partielle dont il manque les parties postérieures aux P2, TM 247-01-02 : fragment de corps mandibulaire droit portant la suite P3-M3).
- quatre dents isolées (TM 266-01-447 : bourgeon de M3 supérieure droite, TM 266-01-448 : I1 supérieure droite, TM 266-02-154-2 : canine inférieure droite, publiées en 2002 et une P3 supérieure droite, TM 266-01-462, publiée en 2005).
- une diaphyse de fémur gauche (TM 266-01-063).[11],[12]
Si il est ainsi possible de reconstituer la tête, il est délicat d'avoir une idée précise du reste du corps.
Caractéristiques morphologiques
Caractères diagnosiques de Sahelanthropus tchadensis
Les caractères diagnosiques du genre et de l’espèce sont les suivants.[2],[10],[13]
Crâne
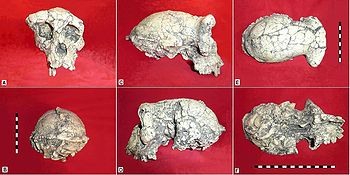 Moulage du crâne holotype non-reconstruit de Sahelanthropus tchadensis TM 266-01-060-1, surnommé Toumaï (orientation approximative selon le plan de Francfort horizontal).
Moulage du crâne holotype non-reconstruit de Sahelanthropus tchadensis TM 266-01-060-1, surnommé Toumaï (orientation approximative selon le plan de Francfort horizontal).
A : vue antérieure
B : vue postérieure
C : vue latérale droite
D : vue latérale gauche
E : vue supérieure
F : vue inférieure
Echelle identique pour toutes les photos : 1 case = 1 cm.en vue antérieure :
- face haute
- épais bourrelet supra-orbitaire (18,2 mm avant reconstruction, 16 mm après), continu (apomorphie supposée)
- large pilier inter-orbitaire
- pas de fosse canine entre I2 et C (apomorphie)
- partie supérieure de la face large par rapport à la partie inférieure qui est étroite dans le plan frontal et courte antéro-postérieurement (apomorphie)
- légère dépression de l’écaille frontale sans former de trigône comme chez Paranthropus
en vue latérale :
- écaille de l’os frontal plate en vue latérale
- pas de sulcus (gouttière ou dépression allongée) derrière le bourrelet supra-orbitaire, à la différence des grands singes.
- trou auditif externe de forme ronde
- crête temporo-nucale composée marquée (importante musculature de la nuque, caractère plésiomorphe)
- prognathisme sub-nasal faible (de l’os maxillaire, en dessous du nez), face relativement droite, orthognathe (apomorphie).
- petite crête sagittale située postérieurement
- plan nucal subhorizontal, relativement plat et relativement long (apomorphie) avec une grande crête occipitale externe.
en vue supérieure :
- boîte crânienne allongée
- constriction post-orbitaire marquée
en vue postérieure :
- processus mastoïdes développés, toujours en relation avec l’importante musculature de la nuque.
en vue inférieure :
- trou occipital (foramen magnum) en position relativement avancée, compatible avec une locomotion bipède (apomorphie).
- forme du trou occipital certainement allongée antéro-postérieurement comme chez les représentants de la lignée humaine (bipèdes) et non-circulaire comme chez les grands singes (l’état de conservation de cette zone ne permet pas d’être absolument affirmatif).
- petits condyles occipitaux
- cavité glénoïde de l'os temporal profonde avec un grand processus post-glénoïde (articulation temporo-mandibulaire puissante)
La capacité crânienne, estimée à partir d’une reconstruction virtuelle, est comprise entre 360 et 370 cm3, donc dans la variabilité de celle des chimpanzés.
Mandibule
- arcade dentaire upsiloïde (en forme de U, plésiomorphie), étroite et relativement petite (apomorphie).
- corps mandibulaire robuste et court supéro-inférieurement
- large sulcus (sillon, dépression allongée) extra-molaire (à l’extérieur des molaires)
- symphyse relativement verticale avec un faible torus transverse (apomorphie)
Dents
Formule dentaire mâchoire supérieure 3 2 1 2 2 1 2 3 3 2 1 2 2 1 2 3 mâchoire inférieure Total: 32 Dentition permanente humaine - incisives relativement petites (apomorphie)
- incisives latérales supérieures situées de part et d’autre du bord latéral de l’ouverture nasale, comme chez les chimpanzés (plésiomorphie).
- canines à couronne petite relativement à celles des grands singes, coniques et usées à l’apex, ce qui indique un complexe C-P3 non-tranchant, un caractère dérivé propre aux hominines. Elles ont en revanche une longue racine (caractère plésiomorphe).
- pas de diastème C-P3 sur la mandibule (différence d’avec les grands singes, caractère supposé apomorphe)
- dents jugales plus petites que celles des australopithèques
- épaisseur de l’émail des dents jugales (prémolaires et molaires) intermédiaire entre Pan (chimpanzés et bonobos) et Australopithecus (caractère supposé apomorphe).
- prémolaires inférieures et supérieures à deux racines et 3 canaux pulpaires séparés (plésiomorphie, caractère primitif supposé commun au rameau Homo/chimpanzé)
- molaires à cuspides relativement bas et arrondis et à face linguale convexe (apomorphie)
- M3 supérieure triangulaire et M3 inférieure rectangulaire et arrondie distalement
Restes post-crâniens
À ce jour, la MPFT ne déclare pas avoir trouvé de reste post-crânien. La bipédie est supposée d’après la base du crâne, mais non confirmée.
Cependant, Beauvilain, l'un des co-auteurs de la mise au jour de Toumaï, déclarait depuis plusieurs années sur un site internet qu'il existe 52 restes post-crâniens trouvés sur le site TM 266, dont un fémur gauche d'hominidé.[14] La détermination zoologique de tous ces restes n'a pu être effectuée sur le terrain. 36 d'entre eux sont des os longs (tibias, fémurs, humérus, cubitus...), entiers ou fragmentaires. En conséquence, Beauvilain publie en mai 2009 un article scientifique comprenant la photographie de ce fémur au moment de sa découverte le 19 juillet 2001 à proximité immédiate du crâne de Toumaï.[15]
Extrapolations à partir des caractères observés
Les mensurations des restes crâniens et dentaires récoltés permettent d'estimer que Sahelanthropus tchadensis mesurait 105 à 120 cm, soit une taille semblable à celle du chimpanzé commun Pan troglodytes.[3] ,[16] Les caractères dentaires suggèrent une alimentation plus variée que celle des grands singes, principalement herbivores et frugivores.
Pour ses inventeurs, le bourrelet supra-orbitaire, par sa très forte épaisseur (18,2 mm avant reconstruction) suggère que Toumaï serait un mâle[2]. Il est toutefois impossible d'être affirmatif sur ce point : juger de la variabilité d'un caractère, en l'occurrence un dimorphisme sexuel, en l'absence d'un échantillon suffisant d'individus pour l'espèce est scientifiquement impossible.[17] Paléontologue spécialiste des hominoïdes et hominines miocènes et pliocènes, Brigitte Senut note que « le bourrelet supra-orbitaire n'est pas un caractère classique de dimorphisme sexuel. »[17]
Une épaisseur de 18,2 mm le place en tout cas au-dessus de tout ce qui a pu être mesuré chez les chimpanzés et les gorilles femelles et même mâles. Elle le distingue aussi nettement des australopithèques.[2]
Position phylogénétique
Conclusions des découvreurs : Sahelanthropus est un hominine
Sahelanthropus tchadensis possède différents caractères dérivés propres aux hominines comme une canine à petite couronne et large tubercule apical, usée apicalement, qui indique un complexe C-P3 inférieure non-tranchant, et une épaisseur de l’émail des dents jugales intermédiaire entre celle de Pan et celle des Australopithèques.[2],[3] Différents caractères sont similaires aux hominines plus récents tels que Kenyanthropus et Homo : morphologie de la base du crâne (longue, à orientation horizontale et trou occipital en position antérieure), face (prognathisme subnasal réduit, pas de diastème entre C et P/3, épais bourrelet supraorbitaire). Ces caractères anatomiques indiquent, selon les auteurs, son appartenance à la lignée des hominines. La présence d'un seul de ces caractères dérivés propre aux hominines suffit d'ailleurs à placer Sahelanthropus tchadensis parmi ces derniers, comme le précisent les auteurs.
Sahelanthropus tchadensis présente aussi une série de caractères plésiomorphes (ancestraux) comme une petite capacité crânienne, un os basioccipital en forme de triangle tronqué et la partie pétreuse de l’os temporal orientée à 60° par rapport à la ligne bicarotidienne (ligne parallèle au plan frontal).
Cette mosaïque de caractères dérivés et primitifs indique pour les auteurs que Sahelanthropus tchadensis occupe une position phylogénétique d’hominine strict, proche de la divergence homme-chimpanzé. Le rattachement de Toumaï à la branche des hominines ne fait donc pas de doute pour l'équipe franco-tchadienne qui l'a découvert, ainsi que pour une partie de la communauté scientifique.
La diagnose différentielle établie par les découvreurs montre également que Sahelanthropus tchadensis diffère notablement de tous les primates hominoïdes connus du Miocène : Samburupithecus, Ouranopithecus, Sivapithecus et Dryopithecus, qui ne présentent aucun des caractères dérivés propres à la lignée humaine relevés chez Sahelanthropus.
Suite à une contestation du statut d’hominine de Sahelanthropus tchadensis par un groupe d’auteurs français et anglo-saxons en 2002 dans Nature, Brunet répond, de manière préliminaire, à ces objections dans la même revue.[18] Il note que tous les caractères n'évoluent pas à la même vitesse et que les caractères relevés par ses détracteurs sont des caractères ancestraux hérités qui montrent, comme il l'a déjà énoncé, la proximité du nouvel hominine de la divergence homme-chimpanzés, survenue nécessairement avant 7 millions d'années. Il note en outre que les détracteurs de son hypothèse n'ont pu réfuter aucun des arguments présentés, permettant de classer les restes à l'intérieur du clade des hominines. Constatant le manque d'appui scientifique des propos de ses détracteurs, Brunet juge leur argumentaire « désinvolte ».[18] Il met alors au défi ses détracteurs de trouver un seul caractère dérivé qui soit partagé par Sahelanthropus et un grand singe, seule condition suffisante et scientifiquement acceptable pour reclasser les fossiles parmi les ancêtres directs des grands singes.
De manière plus formelle, les membres de la MPFT répondent à leurs détracteurs par deux nouvelles études publiées en 2005 dans Nature[10],[13] et par un article de synthèse publié la même année dans La Recherche.[19]
La première étude[10] correspond à la publication de trois nouveaux restes (deux fragments de mandibule et une P3 supérieure droite). Elle confirme les caractères décrits à partir des premiers restes publiés et montre la présence de nouveaux caractères dérivés d'hominines : symphyse relativement verticale avec un faible torus transverse.[10]
La deuxième étude[13] se base sur une reconstruction virtuelle tridimensionnelle du crâne holotype dont la méthode est éprouvée scientifiquement (les premiers résultats publiés en 2002 se basaient sur une reconstruction préliminaire). Cette reconstruction correspond à la moyenne de quatre reconstructions obtenues selon deux protocoles menés séparément par des personnes différentes. Cette reconstruction finale a ensuite été confrontée à trois tests indépendants. Elle montre notamment que l’angle entre le plan du foramen magnum et celui des orbites (angle FM-OP) est égal à 95°. Cette valeur est différente de ce qui existe chez Pan troglodytes (63,7 ± 6,2°) et se rapproche de ce qui existe chez l’homme actuel (103,2 ± 6,9°) et chez des hominines bipèdes tels que Australopithecus afarensis et Australopithecus africanus (environ 80°). Cet angle est, selon les auteurs, directement en relation avec la posture de locomotion sachant que, chez tous les primates, le plan des orbites est à peu près perpendiculaire au sol et le plan du trou occipital perpendiculaire à la première vertèbre cervicale. C'est sur la base de cet angle que Brunet affirme : « La probabilité que Sahelanthropus était bipède est plus grande que celle qu'il ne l'était pas. »[19]
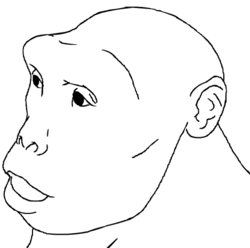 Croquis de reconstitution de Sahelanthropus tchadensis, dans une position droite.
Croquis de reconstitution de Sahelanthropus tchadensis, dans une position droite.Les auteurs ajoutent à cet argument une démonstration de l'écart morphologique existant entre le crâne de Toumaï virtuellement reconstruit et celui d'un chimpanzé ou d'un gorille. Reconstruire le crâne selon un modèle de crâne de chimpanzé ou de gorille aboutit tout simplement à une perte de son intégrité anatomique, des disjonctions apparaissant.[13] Les conclusions sont sans appel pour les auteurs : Sahelanthropus tchadensis a de très fortes chances d’être bipède et s’inscrit dans la lignée humaine. D'autres arguments viennent appuyer la thèse d’une locomotion bipède :
- plan nucal plat orienté à 36° par rapport au plan de Francfort, ce qui entre dans la variabilité du genre Homo et des australopithèques mais pas de Pan ;
- crête nucale présentant une lèvre inférieure, caractère présent chez d’autres hominines bipèdes et absent chez Pan et Gorilla.[13]
Sur la base de l'association originale de caractères primitifs et dérivés, Brunet propose l'existence d'un « nouveau grade d'évolution humaine », antérieur aux grades australopithèque et Homo, qui serait caractérisé notamment par un petit cerveau et la perte de la fonction aiguisoir du complexe C-P3.[3],[19] Pour Brunet, « ce grade aurait pu précéder ou bien être associé à l'acquisition d'une bipédie primitive », attestée avec certitude chez Orrorin tugenensis.[3]
En 2006, les membres de la MPFT publient dans les Proceedings of the National Academy of Sciences[20] une étude morphologique géométrique dont le but est de déterminer plus précisément les affinités morphologiques du crâne TM 266 avec des crânes de chimpanzés, de gorilles et de quelques hominines pliocènes et pléistocènes (échantillon d’australopithèques, Homo habilis, Homo sapiens). Le principe consiste en une superposition procuste des crânes grâce au logiciel MORPHOLOGIKA après avoir enregistré les références d'un certain nombre de points. Cette superposition permet de comparer des conformations 2D ou 3D en éliminant les différences liées à la taille. Une analyse en composantes principales (ACP) montre tout d'abord que le crâne se classe clairement parmi les australopithèques. Par l'analyse procustéenne, les auteurs arrivent à la conclusion qu’en plus de s’inscrire dans le clade des hominines, Toumaï se rapproche en apparence au moins, par son faible prognathisme et la projection relative du neurocrâne au-dessus de la face, plus de KNM-ER 1813 (Homo habilis) que des australopithèques[20]. Ils complètent cette étude par une liste de données comparatives pour 27 caractères crâniens qui révèlent dans quelles mesures Toumaï se rapproche des chimpanzés, des gorilles et des hominines pliocènes et pléistocènes sélectionnés (Australopithecus afarensis, Australopithecus africanus, Australopithecus boisei, Homo habilis, Homo sapiens). Les caractères rapprochant Toumaï des grands singes sont toujours interprétés en termes de plésiomorphies. Les caractères plus proches des hominines ou partagés avec ces derniers sont nombreux et toujours localisés au niveau de la face et de la base du crâne. L’ensemble de ces données leur permet de proposer, de manière préliminaire, un cladogramme construit par la méthode du neighbour joining, dans lequel Sahelanthropus appartient à un clade postérieur à la divergence homme-chimpanzé, groupe frère de tous les autres hominines, auquel pourraient aussi appartenir Orrorin tugenensis et Ardipithecus.[20]
Des contestations : Sahelanthropus est un singe
Cependant, certains scientifiques, comme Milford Wolpoff, Brigitte Senut, Martin Pickford et John Hawks, dont les deuxième et troisième sont co-découvreurs d’Orrorin tugenensis, un autre hominine de la fin du Miocène, suggèrent que Sahelanthropus tchadensis ne serait pas un ancêtre de l'homme mais un ancêtre des grands singes actuels.[21]
En 2002, dans la revue Nature, ils proposent de renommer la découverte de Michel Brunet, « Sahelpithecus », la faisant passer du statut de pré-humain à celui de pré-singe. Ils soulignent sa faible capacité crânienne, la racine de la canine inférieure trop longue, ses orbites sub-carrées rappelant celles des grands singes actuels et du Miocène, le système de crêtes et la surface fortement inclinée à l'arrière du crâne qui rappellerait celle des grands singes quadrupèdes (ils estiment en effet l'orientation du plan nucal à environ 55° par rapport au plan de Francfort). Ils proposent donc de reclasser Toumaï en paléo-gorille et ajoutent que l'on s'est trompé sur son sexe : Toumaï serait une femelle, ce qui expliquerait son faible prognathisme et la petite hauteur des canines.
Ce groupe de spécialistes formule l'idée selon laquelle Brunet et son équipe seraient en train de répéter avec Sahelanthropus une erreur bien connue de l'histoire de la paléoanthropologie, qui avait déjà été commise à propos du Kényapithèque et du Ramapithèque, que l'on avait eux aussi érigés en ancêtres des hominines avant qu'ils ne soient destitués « pour des raisons sexuelles » (il s'agissait en fait de femelles de grands singes miocènes).[21] Brigitte Senut ajoute qu'« en outre, les simiiformes arboricoles ont une tendance à présenter des faces plus plates que les terrestres ».[17]
L'absence d'autres éléments de squelette rend l'analyse de Brunet et ses collaborateurs sujette à controverse, particulièrement au sujet du mode de locomotion, selon eux. Ils interprètent la position avancée du trou occipital et les caractéristiques faciales comme les conséquences d'« une adaptation biomécanique ».[21] Ils appuient ce jugement en citant un certain nombre d'études montrant que « trou occipital antérieur n'est pas synonyme exclusif de bipédie. »[21],[17] Les tenants de cette hypothèse précisent que si Toumaï était l'ancêtre d'un singe, cette découverte resterait un événement exceptionnel d’un point de vue scientifique puisque la paléontologie des grands singes est encore quasiment inconnue.
Suite aux arguments publiés dans Nature en 2005 par Zollikoffer et ses collaborateurs et repris par Brunet et Allemand dans La Recherche en 2005 concernant la posture de locomotion de Toumaï à partir de la mesure de l’angle FM-OP sur le crâne virtuellement reconstruit, deux nouvelles publications réévaluent scientifiquement et infirment les assertions de la MPFT sur lesquelles repose le classement de Sahelanthropus parmi les hominines.
La première, de Martin Pickford, publiée en 2005 dans la revue Anthropologie. Partant d’un constat de contradictions flagrantes, internes à ces publications de la MPFT, entre valeurs données dans le texte et valeurs mesurables sur les figures, Martin Pickford se propose de réévaluer l’angle FM-OP chez le chimpanzé, Homo sapiens et également chez les gorilles, à partir de larges échantillons et selon un protocole qu’il explique en détail.[22]
L’auteur trouve des valeurs en contradiction avec celles publiées par la MPFT. Pour le chimpanzé, il évalue cet angle à 69° en moyenne, avec une amplitude de 60 à 78°, alors que la MPFT l’évalue à 63,7 ± 6,2°[13] ou 65°[19] en moyenne (alors que l’exemple figuré dans les publications de la MPFT montre un angle de 57°, donc plus aigu que la limite inférieure fournie dans le texte, égale à 57,5°).
Ce désaccord est lourd de conséquences en termes d’implications méthodologiques, car si l’évaluation de Pickford est juste, il y a de grandes chances pour que le domaine de variation de cet angle pour le chimpanzé recouvre celui des australopithèques, pour lesquels l’angle fourni par la MPFT (81° chez Australopithecus africanus) est un angle moyen pour ce groupe. L’angle FM-OP pour ces espèces n’étant pas distinguable sans ambiguïté et étant admis que les chimpanzés ont une posture pronograde alors que celle des australopithèques est orthograde (fait appuyé par les nombreuses études de restes post-crâniens), il devient alors impossible de déduire la posture de locomotion à partir de cet angle.[22] Cet angle étant un mauvais indicateur postural pour ces groupes, sa capacité à indiquer la posture de Sahelanthropus, même si l’angle FM-OP de ce dernier fourni par la MPFT (95° dans le texte, 96° sur la figure[13],[19]) est proche de la limite inférieure de variation de cet angle pour Homo sapiens, s’en trouve fortement remise en cause. Par ailleurs le plan orbitaire fourni par la MPFT pour Sahelanthropus est comparable à celui des chimpanzés, gorilles et australopithèques, alors qu’étrangement le plan du foramen magnum est identique à celui d’Homo sapiens, ce qui en soi et d’un point de vue de l’évolution, constitue une incohérence. On peut en conséquence légitimement supposer une mauvaise évaluation du plan du foramen magnum de Toumaï découlant peut-être d’une mauvaise reconstruction du crâne, ayant volontairement ou involontairement, donné une orientation trop horizontale à ce plan (par abaissement du neurocrâne par rapport à la face) et conséquemment augmenté le fameux angle FM-OP.[22]
En conséquence de toutes ces remarques, M. Pickford considère qu'il est prématuré de déduire une posture orthograde et une locomotion bipède pour Sahelanthropus sur la base de son angle FM-OP reconstruit. Ceci renforce l’idée de sa non appartenance aux hominines.[22]
La deuxième publication, de Wolpoff, Hawks, Senut, Pickford et Ahern parue en 2006 dans la revue PaleoAnthropology, postérieurement à la dernière publication de la MPFT,[20] réévalue scientifiquement l’ensemble des piliers soutenant l’hypothèse du rattachement de Toumaï au rameau humain : taille de la canine, mode et amplitude de l’usure de celle-ci, plan nucal et position de l’inion, angle entre plan du foramen magnum et plan des orbites, en estimant en premier lieu son âge et son sexe qui sont des paramètres influant sur l’évaluation et l’interprétation des caractères précédents.[23]
Ils estiment que le crâne TM 266 correspond à celui d’un individu de plus ou moins 11 ans et dont le sexe est indéterminable étant donnés les indices équivoques fournis par la taille de la canine et l’épaisseur du bourrelet sus-orbitaire (16 mm après reconstruction),[23] contrairement à Brunet et al. 2002 qui pense qu'il s'agit d'un mâle d’environ 20 ans.
La taille relative et absolue de la canine est proche de la moyenne connue pour les singes miocènes. Son type d’usure se rencontre chez de nombreux autres singes miocènes, particulièrement Ouranopithecus et Gigantopithecus. L’apparition d’un complexe C-P3 moins tranchant, déjà présent chez les grands singes miocènes, doit donc être considérée comme une homoplasie : une adaptation à une mastication puissante. La forte amplitude de cette usure, inhabituelle pour son âge, laisse supposer également une cause masticatrice et ne peut induire des conclusions phylogénétiques.[23] Concernant le deuxième pilier soutenant l’appartenance aux hominines, à savoir les preuves crâniennes d’une posture droite et d’une locomotion bipède, les auteurs montrent que[23] :
- la forme allongée et plate, la largeur, l’angulation et de nombreux détails du plan nucal sont similaires à ce qui s'observe chez les singes,
- la lèvre inférieure de la crête nucale correspond au fort développement du ligamentum nucale en relation avec une puissante musculature,
- la longueur relative du plan nucal (longueur entre inion et opisthion, par rapport à la capacité crânienne ici) excède largement celle de tous les hominidés connus,
- l’inclinaison du plan nucal par rapport au plan de Francfort (36° après reconstruction, environ 55° avant) implique, dans tous les cas, une direction des muscles nucaux semblable à celle observée chez le chimpanzé et donc une posture similaire,
- le plan nucal s’étend au-dessus du plan de Francfort jusqu’au sommet de l’arrière-crâne, ce qui contraste avec tous les hominines, pour lesquels le haut du plan nucal (inion) est situé dans ce plan ou à proximité de celui-ci. L’indice de la hauteur du plan nucal de Le Gros Clark (1950), c’est-à-dire la hauteur du sommet du plan nucal (inion) au-dessus du plan de Francfort rapportée à la hauteur maximale du crâne au-dessus de ce même plan et multipliée par 100, calculé à partir de la reconstruction (23) est très au-dessus des valeurs de cet indice pour les hominines (maximum 14 chez WT 17000, Paranthropus aethiopicus) et correspond aux valeurs minima calculées chez les chimpanzés (23) et les bonobos (21).
- l’angle FM-OP n’est pas un bon indicateur de posture étant donnés les recouvrements observés dans la distribution des valeurs pour 63 spécimens de chimpanzés (41°-80°) et 4 australopithèques (64°-81°), deux groupes de primates dont on sait qu’ils ont pourtant des postures et modes locomoteurs différents. Si cet angle n’est pas un bon indicateur postural chez les chimpanzés et les australopithèques, il est impossible qu’il puisse l’être pour Sahelanthropus, espèce supposée proche de la divergence menant à ces deux groupes.
Par ailleurs, les auteurs insistent sur le désaccord de l’hypothèse « hominine » avec les estimations de la date de dichotomie chimpanzés-hominines par la biologie moléculaire.
En conséquence, pour eux, le crâne TM 266 montre des contrastes profonds avec tous les hominines connus et cela suffit pour l’exclure de leur filiation. C’est cette exclusion, et non une quelconque série de caractères synapomorphes, qui assigne à Sahelanthropus une place parmi les singes au sens large, soit antérieure, soit postérieure à la dichotomie hominines-chimpanzés.[23]
Des soutiens à l'interprétation comme hominine
 Yves Coppens, l'un des auteurs soutenant l'interprétation de Sahelanthropus tchadensis comme hominine.
Yves Coppens, l'un des auteurs soutenant l'interprétation de Sahelanthropus tchadensis comme hominine.Yves Coppens, qui a été co-signataire de plusieurs publications[2],[3] dont la première de Sahelanthropus tchadensis, reconnaît avoir été surpris par la position avancée du trou occipital. Il « pencherait volontiers » pour placer ce fossile, malgré ses allures d’ « humain par devant » (face moderne) et de « grand singe par derrière » (système de crêtes), après la dichotomie et du côté pré-humain.[24]
David Pilbeam et Daniel E. Lieberman, paléoanthropologues renommés de l'université Harvard sont, comme Yves Coppens, co-signataires de plusieurs articles de Brunet ou de ses collaborateurs au sujet de Sahelanthropus tchadensis.[2],[3],[10],[13] Le premier est, comme Yves Coppens, membre de la MPFT. Le second avait déclaré, avant la première publication, que cette découverte ferait « l'effet d'une bombe nucléaire dans le milieu de la paléoanthropologie ».[25] Ce même auteur déclarait le 14 juillet 2002, quelques jours après la première publication de Brunet et ses collaborateurs dans Nature, son agrément à la thèse d’un hominine bipède : « […] and it looks as if it might have walked upright. These features are typical of later hominids rather than apes. »[26]
William Kimbel, directeur de l’Institute of Human Origins de l’université de l’Arizona, dans un article critique paru en 2004 au sujet d’un ouvrage de Jonathan Kingdon, se positionne de la même façon « […] the 6- to 7-million-year-old Sahelanthropus tchadensis fossil from Chad, whose canine-tooth and skull-base characteristics appear to place it on our side of the chimp-human split […] » et va même jusqu’à juger Sahelanthropus plus convaincant qu’Orrorin tugenensis dans son aptitude à la bipédie et son appartenance aux hominines.[27] De même David Begun marque en 2004 son soutien à la thèse d’un hominine bipède.[28]
Yohannes Haile-Selassie, Gen Suwa et Tim White, partisans de la même thèse, vont plus loin en suggérant, sur la base des dents, que Sahelanthropus et Ardipithecus pourraient constituer un même taxon.[29]
Une étude scientifique approfondie et indépendante, de Strait et Grine, parue en 2004, donne crédit à l'interprétation phylogénétique de la MPFT. Dans cette étude cladistique basée sur la comparaison de 198 caractères crânio-dentaires chez plusieurs fossiles d'hominines reconnus et supposés et chez les hominoïdes actuels (gibbon, orang-outan, gorille, chimpanzé), les auteurs testent entre autres hypothèses phylogénétiques concernant divers fossiles, l'hypothèse émise dès 2002 par la MPFT[2] (et soutenue dans les publications postérieures[3],[20]) selon laquelle Sahelanthropus tchadensis se place après la divergence chimpanzé-hominines et peut être considéré comme groupe frère de tous les autres hominines. Leur analyse est cohérente avec cette interprétation même si la proximité phylogénétique entre Sahelanthropus et Ardipithecus est jugée relativement instable (deux arbres obtenus donnent des résultats en désaccord avec les autres sur ce point).[30]
Objections à la thèse d’un hominine
Caractères convergents au sein d'une radiation adaptative
Bernard Wood, en 2002 dans Nature, reconnaît à Sahelanthropus ses caractères apparents d’hominine : « What is remarkable about the chimp-sized cranium TM 266-01-060-1 discovered by Brunet et al. is its mosaic nature. »[31] Cependant accepter Toumaï en ancêtre du rameau humain reviendrait à concevoir la phylogénie des premiers hominines et des singes miocènes selon un modèle linéaire (« tidy model »), auquel Wood n’adhère pas puisqu’il pense qu’au contraire une grande diversité taxinomique existait entre 5 et 7 millions d’années (« untidy model » ou « bushy model »). Ce deuxième modèle prévoit la possibilité de nombreuses homoplasies et le bourrelet sus-orbitaire de Toumaï par exemple pourrait en être une. Wood pense que Sahelanthropus est un candidat à l’ancêtre des hominines, mais qu’il est impossible de prouver qu’il en est un.[31] Il conclut en exprimant l’idée selon laquelle Sahelanthropus constitue la preuve d’une diversité taxinomique incluant l’ancêtre commun aux panines (chimpanzés actuels et fossiles) et aux hominines (humains actuels et fossiles), diversité sûrement comparable à celle de la faune d’invertébrés de Burgess il y a 500 millions d’années : « My prediction is that S. tchadensis is just the tip of an iceberg of taxonomic diversity during hominid evolution 5-7 million years ago. Its potentially close relationship with our own, hominid, twig of the tree of life is surely important. More notably, however, I think it will prove to be telling evidence of the adaptative radiation of fossil ape-like creatures that included the common ancestor of modern humans and chimpanzees. »[31]
Sahelanthropus tchadensis en attente de classement
Dans un article intitulé « Genres de la lignée humaine » publié en 2003, Camilo J. Cela-Conde et Francisco J. Ayala, après avoir défini ce qu’ils entendaient par Hominidé, proposent l’existence de seulement quatre genres pour la lignée humaine correspondant à quatre zones d'adaptation : Praeanthropus dont l’espèce souche serait Praeanthropus tugenensis (Orrorin tugenensis), Ardipithecus, Australopithecus dont l’espèce souche serait Australopithecus africanus et Homo dont l’espèce souche serait Homo platyops (Kenyanthropus platyops), auxquels ils ajoutent un genre incertae sedis, c’est-à-dire de statut incertain, pour Sahelanthropus tchadensis.[32] Ils considèrent en effet que l’absence de restes post-crâniens, la proximité d’avec la divergence panines-hominines et la localisation géographique de Sahelanthropus justifient ce classement provisoire. Ils précisent cependant qu’ « il paraît pour le moment impossible de déterminer avec certitude le classement des nouveaux spécimens du Miocène »[32] (Orrorin tugenensis, Ardipithecus ramidus s. l. et Sahelanthropus tchadensis).
Une question méthodologique
Pascal Picq rappelle combien il est difficile de discriminer un caractère dérivé d’un caractère primitif lorsqu’on a peu de fossiles, ce qui est le cas pour la période allant de 12 à 7 Ma, correspondant à « des origines dans un désert de fossiles ».[33] Si d'après les travaux les plus récents de la cladistique ou systématique phylogénétique, les chimpanzés et le rameau humain partagent un dernier ancêtre commun exclusif, on n'en connaît pas les caractéristiques.[34] Il souligne ainsi un important problème méthodologique de la discipline, critique qui ne touche pas seulement Sahelanthropus tchadensis mais aussi tous les autres fossiles.
Les paléoanthropologues ont souvent confondu caractères archaïques de la lignée humaine avec caractères actuels des grands singes et ont analysé les fossiles selon deux pôles, « ape-like » et « human-like », pour juger de la position phylogénétique alors que ces deux pôles correspondent à des états dérivés et que les caractères ancestraux communs restent inconnus. Personne aujourd’hui n’est en mesure de savoir quels étaient les caractères des individus situés juste avant la dichotomie homme-chimpanzé. Il est donc délicat de juger de l’état dérivé ou primitif des caractères observés sur Sahelanthropus et d’en déduire sa position phylogénétique et son statut taxinomique. La bipédie, que l’on tient habituellement pour un caractère dérivé propre à la lignée humaine, peut aussi bien être un caractère ancestral.
La cladistique et la systématique moléculaire, qui reposent sur le concept de l'horloge moléculaire, permettent d'envisager une date de divergence comprise entre 1 et 15 millions d'années, avec un moyenne autour de 7 Ma.[35]. Sahelanthropus peut, selon Pascal Picq, se situer avant ou après cette dichotomie.[24]
« Avec Toumaï et Orrorin, on flirte avec le dernier ancêtre commun.[...] La question est de savoir lequel est le plus proche et si l'un ou l'autre se situe avant ou après la dichotomie. Au rythme actuel des découvertes, on peut s'attendre à bien des surprises.[36] »Conclusions provisoires
La bipédie de Sahelanthropus tchadensis est très probable pour ses découvreurs. En effet, le trou occipital correspond à celui d'une colonne vertébrale redressée, ce qui permet d'inférer une locomotion bipède.[2],[13] Cependant l'absence actuelle de restes autres que le crâne, des mandibules fragmentaires et des dents isolées, ne permet pas encore d'être catégorique sur ce point ; la bipédie n'est pas confirmable tant que l'analyse d'un os du membre inférieur ou du bassin n'est pas effectuée.[2],< ref name="Brunet et al., (2004)"/>,[13],[21]
Une partie de la communauté scientifique s'accorde à reconnaître le statut d'hominine à Sahelanthropus tchadensis ainsi que son aptitude à la bipédie. Cependant, l'hypothèse selon laquelle Sahelanthropus serait un singe, situé avant ou après la divergence panines-hominines, a très tôt été émise. La position phylogénétique de Sahelanthropus a fait et fait encore l'objet d'une controverse entre spécialistes. Cette controverse a plusieurs causes, outre les divergences d'interprétation fonctionnelle des caractères :
- son statut d'hominine corrélé à son ancienneté est en désaccord avec les données de la génétique, qui placent la séparation homme-chimpanzé au plus entre 5 et 6,5 millions d'années, selon les études, même les plus récentes.[37],[23],[32]
- il existe un manque de données sur les caractéristiques du dernier ancêtre commun aux chimpanzés et aux hominines, et il existe par conséquent un biais méthodologique lié à la validité des caractères considérés comme propres à la lignée humaine.[38],[39],[24]
- les spécialistes se disputent l'honneur de détenir le plus vieux représentant de la lignée conduisant à l'homme moderne,[32] privilège qui revenait avant la découverte de Toumaï, mais de manière tout aussi controversée, à Orrorin tugenensis, surnommé « Millenium ancestor ».
- la MPFT ne permet pas la consultation des fossiles originaux par des équipes extérieures.
- la MPFT n’a pas reconnu avant 2009 être en possession d’un fémur pourtant mis au jour le 19 juillet 2001 à proximité immédiate du crâne de Sahelanthropus tchadensis.
Il s'agit en fait d'un vaste débat naissant sur les plus anciens hominines, voire sur l'ancêtre commun avec les chimpanzés et pré-chimpanzés, qui concerne d'autres fossiles à la charnière Miocène-Pliocène (Orrorin tugenensis, Ardipithecus kadabba). Ces espèces qui présentent « toutes des caractéristiques primitives et quelques unes évoluées […] posent la question de savoir comment les Homininés sont apparus, et corrélativement quels caractères peuvent être considérés comme diagnostiques de cette lignée à son origine. »[38]
En guise de conclusion provisoire, le professeur de paléoanthropologie José Braga, à l'université Paul Sabatier de Toulouse, écrit à propos des restes du Miocène supérieur et du Pliocène inférieur, que sont Sahelanthropus tchadensis, Orrorin tugenensis et Ardipithecus ramidus s. l., « Le statut d'hominidé des fossiles, encore peu nombreux, découverts dans ces périodes chronologiques, reste discuté, à des degrés différents, notamment, selon leur état de préservation. »[40] En suivant l'avis général émanant de la communauté scientifique, nous pouvons donc dire de Sahelanthropus tchadensis qu'« il s'agit actuellement de l'un des trois candidats au titre d'ancêtre de la lignée humaine. »[38]
Environnement
 Un delta de fleuve endoréique actuel, le delta de l'Okavango
Un delta de fleuve endoréique actuel, le delta de l'OkavangoL’étude géologique et paléontologique du site de Toros-Menalla a été effectuée par Patrick Vignaud et ses collaborateurs afin de dater le site et de reconstituer l’environnement.[41],[3] L’unité sédimentaire à anthracothères ("Anthracotheriid Unit" : AU), qui renferme Sahelanthropus tchadensis, correspond à un environnement péri-lacustre (grès) régulièrement soumis aux inondations dues aux variations de la ligne de rivage d'un paléo-lac Tchad, beaucoup plus étendu que l'actuel.
La flore et surtout la faune de vertébrés retrouvées dans cette unité sédimentaire indiquent des paysages mosaïques, depuis la forêt galerie en bordure de lac (plante légumineuse du type liane, colobes, python) jusqu’à la savane arborée et aussi très ouverte (proboscidiens, girafe, équidés, bovidés, hyénidés, et le grand félidé Machairodus) en bordure de désert. Des formes aquatiques et amphibies indiquent la présence d’eau avec certitude (dix taxons de poissons dulçaquicoles, deux espèces de crocodiles et une nouvelle espèce de gavial, tortues, anthracothères, hippopotame, loutre…).
Parmi les poissons retrouvés, on distingue des formes d'eaux profondes et bien oxygénées caractéristiques d'un lac, des formes adaptées à des milieux marécageux pauvres en oxygène et des formes adaptées à la fois à des milieux marécageux et à des eaux plus turbides (rivière ou fleuve), ce qui implique la présence conjointe d'un lac et d'un fleuve, qui devait certainement l'alimenter.
Il est impossible pour l’instant de déterminer plus précisément parmi cette mosaïque d’environnements péri-lacustres quel était l’environnement précis de Toumaï autour d'un lac endoréique alimenté par plusieurs fleuves. Néanmoins, par la diversité de leurs paysages (de la forêt galerie à la prairie herbeuse en bordure de désert) et de leurs faunes, le lac Tchad actuel et le delta du Chari, voire le delta actuel de l'Okavango, dans le Kalahari central (Botswana) doivent être comparables sans avoir la grande profondeur du paléo-lac.
Datation
Une datation relative, biochronologique
Des méthodes de datation relative biochronologiques ont permis d'estimer l’âge de Toumaï à environ 7 millions d'années, en se fondant sur le degré d'évolution des mammifères, particulièrement des proboscidiens (Anancus kenyensis et Loxodonta sp.) et des suiformes (le suidé Nyanzachoerus syrticus), présents à ses côtés, par comparaison avec d'autres faunes africaines similaires dont l'âge radiochronologique était connu.[42] En effet, il apparaît que la faune du site TM 266 se rapproche plus de celle des niveaux de base de la formation de Nawata à Lothagam (datés autour de 7 millions d’années) que de celle de la formation de Lukeino (celle qui a livré Orrorin tugenensis, datée autour 6 millions d’années).[41]
Une tentative de datation absolue, radiométrique
Une série de dates absolues a été publiée en mars 2008 par une équipe française pour les niveaux de sédiments qui renfermaient le fossile. Ces dates, réalisées grâce à la méthode du béryllium 10 / béryllium 9, ont permis d'établir que l'âge de Toumaï est compris entre 6,8 et 7,2 millions d'années.[37] Toutefois, le crâne de Toumaï n’étant pas en place (in situ) dans le sédiment[43] (noter que les coupes géologiques publiées par la MPFT varient fortement de 2002 à 2008) et a même été déplacé lors d’une vraisemblable inhumation récente,[15] cette méthode ne peut être appliquée.
Implications de la découverte
La fin de l’East Side Story ?
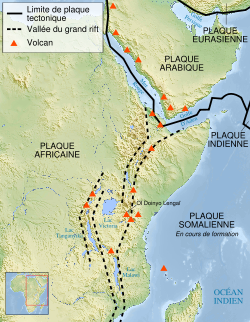 Carte du Rift est-africain.
Carte du Rift est-africain.Dans l'option selon laquelle Sahelanthropus tchadensis est bien un hominine, sa découverte 2600 km à l’ouest de la Rift Valley montre une répartition géographique des représentants miocènes et pliocènes de notre lignée plus large que celle prévue par la théorie de l’East Side Story d’Yves Coppens.[44],[45] avec laquelle s’accordaient encore les découvertes d’Ardipithecus ramidus et d’Orrorin tugenensis.
Yves Coppens reconnaît lui-même que cette découverte, comme celle d’Abel (Australopithecus bahrelghazali), démontre la non-validité de l’East Side Story telle qu’il l’a formulée en 1982. Ce qui ne veut pas dire pour autant, selon Coppens, que la barrière de la Rift Valley n’a pas eu d’importance dans l’histoire évolutive des faunes africaines, primates compris. La considérant comme un filtre, il reste à déterminer selon lui, sa perméabilité différentielle pour chaque groupe et chaque époque.[24]
Le 21 août 2005, lors de sa leçon de clôture au Collège de France, Yves Coppens dit, à propos des fossiles de la lignée humaine, genre Homo exclu : « Ces préhumains se distribuent à travers l'auréole de savanes des tropiques de l'Afrique, du Tchad à l'Éthiopie, au Kenya, à la Tanzanie, au Malawi et à l'Afrique du Sud, entre neuf millions et demi d'années peut-être et un petit peu plus d'un million d'années sans doute. »[46] Autrement dit, si l'aspect éco-évolutionniste de sa théorie n'est pas directement ébranlé par cette découverte en particulier, l'aspect géographique l'est par contre totalement. Pour Coppens, le paradigme a changé.
Notons que si Sahelanthropus est un singe situé avant ou après la divergence des pré-chimpanzés et des pré-humains, son existence en Afrique centrale ne remet nullement en question la théorie de l'East Side Story.
Une remise en question de la date de séparation homme-chimpanzé
La deuxième conséquence majeure de cette découverte est une remise en question de la date de séparation homme/chimpanzé.
Si Sahelanthropus tchadensis est bien un hominine, alors cette séparation doit être au moins antérieure à 7 millions d’années, ce qui contredit la plupart des études moléculaires qui placent celle-ci entre 5 et 7 millions d’années. Toumaï contredit également la récente théorie[47] selon laquelle cette séparation serait complexe et aurait permis des hybridations hominines-panines jusqu’à la séparation définitive de ces deux rameaux il y a moins de 6,3 millions d’années.
Se basant sur la datation absolue de Toumaï, Lebatard et ses collaborateurs[37] considèrent que la séparation hominines/panines a dû intervenir avant 8 millions d’années, ce qui semble s’accorder avec la récente découverte d’un paléo-gorille, Chororapithecus abyssinicus, daté de 10 millions d’années.[48]
L'ancienneté et la position géographique de Sahelanthropus tchadensis ont donc profondément remis en question les idées dominantes concernant l'origine des hominines.
S. tchadensis et les hominines plus récents
Il semble bien établi que la différence d'âge et les différences morphologiques entre Sahelanthropus tchadensis et les Australopithèques, Kenyanthropus et Homo sont importantes et qu'il s'agit de genres différents.[2],[3]
Il reste en revanche difficile de réaliser un arbre phylogénétique. Sans doute faudra-t-il encore de nombreuses découvertes pour permettre de comprendre les relations phylogénétiques liant précisément les genres Homo, Australopithecus et Kenyanthropus avec ce que l'on appelle les « hominidés du Miocène supérieur », de plus de 5,5 millions d'années, que sont Sahelanthropus tchadensis, Orrorin tugenensis et Ardipithecus kadabba.
Succession des différentes espèces d'hominines dans le temps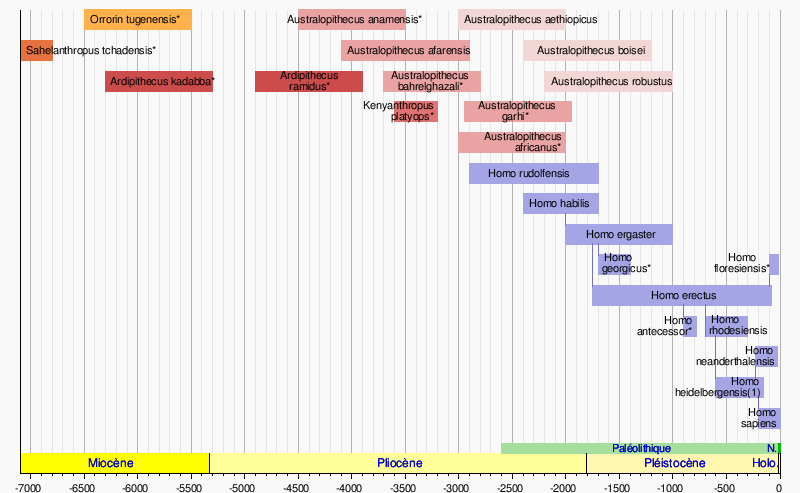
Échelle en milliers d'années Les âges des espèces marquées * sont estimées. - Les traits verticaux symbolisent les possibles lignées (hypothèse "splitter", dite de l'origine unique) (1) ou Homo sapiens archaïque ancien Sources : [2] - [3] - [4] - [5] Sahelanthropus tchadensis et les autres « hominines du Miocène supérieur »
Une question importante reste posée : faut-il regrouper Sahelanthropus tchadensis, Orrorin tugenensis et Ardipithecus kadabba dans un seul genre ?
En 2008, peu d'éléments permettent de répondre. En effet, les restes attribués à ces trois taxons ne se recoupent guère (crâne, mandibules et dents pour Sahelanthropus tchadensis ; dents, fémur, humérus pour Orrorin tugenensis ; dents, bras, phalanges de pied pour Ardipithecus kadabba). Pour Michel Brunet comme pour le découvreur d'Ardipithecus kadabba,[3] l'analyse des dents permet d'avancer que Orrorin et Ardipithecus seraient plus apparentés entre eux que Sahelanthropus ne l'est avec l'un ou l'autre. Les recherches en cours dans les trois pays de découverte (Tchad, Kenya et Éthiopie) pourraient éclaircir cette question dans les années à venir.
Notes et références
- ↑ Dans cet article nous emploierons, en suivant la classification phylogénétique actuelle, le terme hominine (ou hominini en latin) pour désigner les espèces rattachées au rameau conduisant exclusivement à l'homme moderne, postérieures à la divergence d'avec les chimpanzés. Le terme « hominidé » (« hominid » en anglais, « hominidae » en latin) dans le sens de la classification classique (Gray, 1825) désigne ce même clade et c'est en ce sens qu'il est employé par les auteurs cités dans le texte ou en référence du présent article. Le terme homininé, qui au sens de la classification phylogénétique actuelle désigne les tribus hominine et panine, est également employé, par certains auteurs (cités ici), pour désigner uniquement le rameau humain (hominine).
- ↑ a , b , c , d , e , f , g , h , i , j , k , l , m et n Brunet M., Guy F., Pilbeam D., Mackaye H. T., Likius A., Djimdoumalbaye A., Beauvilain A., Blondel C., Bocherens H., Boisserie J.-R., de Bonis L., Coppens Y., Dejax J., Denys C., Duringer P., Eisenmann V., Fanone G., Fronty P., Geraads D., Lehmann T., Lihoreau F., Louchart A., Mahamat A., Merceron G., Mouchelin G., Otero O., Pelaez Campomanes P., Ponce De Leon M., Rage J.-C., Sapanet M., Schuster M., Sudre J., Tassy P., Valentin X., Vignaud P., Viriot L., Zazzo A. et Zollikofer C. P. E., 2002. A new hominid from the Upper Miocene of Chad, Central Africa. Nature, 418 (6894): 145-151.
- ↑ a , b , c , d , e , f , g , h , i , j et k Brunet M., Guy F., Boisserie J.-R., Djimdoumalbaye A., Lehmann T., Lihoreau F., Louchart A., Schuster M., Tafforeau P., Likius A., Mackaye H. T., Blondel C., Bocherens H., De Bonis L., Coppens Y., Denis C., Duringer P., Eisenmann V., Flisch A., Geraads D., Lopez-Martinez N., Otero O., Pelaez Campomanes P., Pilbeam D., Ponce de León M., Vignaud P., Viriot L., Zollikofer C. P. E., 2004. Toumaï, Miocène supérieur du Tchad, le nouveau doyen du rameau humain. C.R. Palevol, 3 (2004): 275-283.
- ↑ Dépêche de l'Associated Press du 12 juillet 2002.
- ↑ Michel Brunet au Collège de France, site du CNRS.
- ↑ La mise au jour d’Abel
- ↑ Brunet M., Beauvilain A., Coppens Y., Heintz E., Moutaye A.H.E. et Pilbeam D., 1995. The first australopithecine 2,500 kilometres west of the Rift Valley (Chad). Nature, 378 (6554): 273-275.
- ↑ La mise au jour de Toumaï.
- ↑ Aujourd'hui disparus, les anthracothères étaient des mammifères omnivores proches de l'actuel hippopotame.
- ↑ a , b , c , d , e et f Brunet M., Guy F., Pilbeam D., Lieberman D. E., Likius A., Mackaye H. T., Ponce de Leon M. S., Zollikofer C. P. E. et Vignaud P., 2005. New material of the earliest hominid from the Upper Miocene of Chad. Nature, 434 (7034): 752-755.
- ↑ Nicolas Constans, 2009. Le fémur de Toumaï ? La Recherche, (432) juillet-août 2009: 18-19.
- ↑ Hawks, 2009. Sahelanthropus: The femur of Toumaï.
- ↑ a , b , c , d , e , f , g , h , i et j Zollikofer C. P. E., Ponce de Leon M. S., Lieberman D. E., Guy F., Pilbeam D., Likius A., Mackaye H. T., Vignaud P. et Brunet M., 2005. Virtual reconstruction of Sahelanthropus tchadensis. Nature, 434 (7034): 755-759.
- ↑ site sur Toumaï
- ↑ a et b Beauvilain A. et Watté J.-P., 2009. Toumaï ("Sahelanthropus tchadensis") a-t-il été inhumé ? Bulletin de la Société géologique de Normandie et des Amis du Museum du Havre, 96 (1): 19-26.
- ↑ dossier de presse du CNRS
- ↑ a , b , c et d Senut B., 2005. Les grands singes miocènes et l'origine des hominidés, in Origine et évolution des populations humaines, Dutour O., Hublin J.-J., Vandermeersch B., (dir.), Comité des Travaux Historiques et Scientifiques, pp. 23-24. ISBN 2-7355-0584-7
- ↑ a et b Brunet M., 2002. Reply to Sahelanthropus or "Sahelpithecus"? Nature, 419 (6907): 582.
- ↑ a , b , c , d et e Brunet M. et Allemand L., 2005. Un nouveau grade d'évolution humaine, Dossier spécial Toumaï. La Recherche, (387), juin 2005: 36-43.
- ↑ a , b , c , d et e Guy F., Lieberman D. E., Pilbeam D., Ponce de Leon M. S., Likius A., Mackaye H. T., Vignaud P., Zollikofer C. P. E. et Brunet M., 2006. Morphological affinities of the Sahelanthropus tchadensis (Late Miocene hominid from Chad) cranium [1] PDF fulltext Supporting Tables, PNAS, 102 (52): 18836-18841.
- ↑ a , b , c , d et e Wolpoff M. H., Senut B., Pickford M. et Hawks J., 2002. Sahelanthropus or "Sahelpithecus"? Nature, 419 (6907): 581-582.
- ↑ a , b , c et d Pickford M., 2005. Orientation of the foramen magnum in late Miocene to extant African apes and hominids. Anthropologie, XLIII (23): 191-198.
- ↑ a , b , c , d , e et f Wolpoff M. H., Hawks J., Senut B., Pickford M. et Ahern J., 2006. An Ape or the Ape : Is the Toumaï Cranium TM 266 a Hominid? PaleoAnthropology, 2006: 36-50.
- ↑ a , b , c et d Toumaï, le trouble-fête ? Page de la cité des sciences, interviews sonores
- ↑ Morin H. 2002. Au Tchad, les restes d'un crâne bouleversent les origines de l'humanité. Le Monde, 11 juillet 2002 copie de l'article
- ↑ Lieberman D. E., 2002. « Upending the Expectations of Science ». The New York Times, 14 juillet 2002, p. 15.
- ↑ Kimbel W. H., 2004. Becoming bipeds : review of "Lowly Origin : Where, When, and Why our Ancestors first Stood Up" by J. Kingdon. American Scientist, 92 (3): 274-276.
- ↑ Begun D. R., 2004. The Earliest Hominins--Is Less More? Science, 303 (5663): 1478-1480.
- ↑ Haile-Selassie Y., Suwa G., White T. D., 2004. Late Miocene Teeth from Middle Awash, Ethiopia, and Early Hominid Dental Evolution. Science, 303 (5663): 1503-1505.
- ↑ Strait D. S., Grine F. E., 2004. Inferring hominoid and early hominid phylogeny using craniodental characters: the role of fossil taxa. Journal of Human Evolution, 47, (6): 399-452.
- ↑ a , b et c Wood B., 2002. Hominid revelations from Chad. Nature, 418 (6894): 133-135.
- ↑ a , b , c et d Cela-Conde C. J. et Ayala F. J., 2003. Genera of the human lineage. PNAS, 100 (13): 7684-7689.
- ↑ Picq P., Au commencement était l'homme. Odile Jacob, 2003, p. 29.
- ↑ Picq P., p. 21.
- ↑ Picq P., p. 28.
- ↑ Picq P., p. 38.
- ↑ a , b et c Lebatard A.-E., Bourles D. L., Duringer P., Jolivet M., Braucher R., Carcaillet J., Schuster M., Arnaud N., Monie P., Lihoreau F., Likius A., Mackaye H. T., Vignaud P. et Brunet M., 2008. Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene hominids from Chad. PNAS 105(9): 3226-3231. PDF fulltext
- ↑ a , b et c Grimaud-Hervé D., Serre F., Bahain J.-J., Nespoulet R., 2005. Histoire d'ancêtres. La grande aventure de la préhistoire. Artcom', Paris, 4e édition revue et augmentée, 135 p., p. 26. ISBN 2-87772-311-9
- ↑ Braga J., 2005. Avant le genre Homo, in Origine et évolution des populations humaines. Dutour O., Hublin J.-J., Vandermeersch B. (dir.), Comité des Travaux Historiques et Scientifiques, pp. 35-65. ISBN 2-7355-0584-7
- ↑ Braga J., p. 39.
- ↑ a et b Vignaud P., Duringer P., Mackaye H. T., Likius A., Blondel C., Boisserie J.-R., de Bonis L., Eisenmann V., Etienne M.-E., Geraads D., Guy F., Lehmann T., Lihoreau F., Lopez- Martinez N., Mourer-Chauvire C., Otero O., Rage J.-C., Schuster M., Viriot L., Zazzo A. et Brunet M., 2002. Geology and paleontology of the Upper Miocene Toros-Menalla hominid locality, Chad. Nature, 418 (6894):152-155.
- ↑ Toumaï, 7 millions d'années, est le « doyen de l'humanité »
- ↑ Beauvilain A., 2008. The contexts of discovery of "Australopithecus bahrelghazali" and of "Sahelanthropus tchadensis" (Toumaï) : unearthed, embedded in sandstone or surface collected ? South african Journal of Science, 104 (3): 165-168.
- ↑ Kortlandt A., 1972. New perspectives on ape and human evolution. Stichting voor Psychobiologie, Amsterdam, 1972
- ↑ Yves Coppens, 1983. Le singe, l’Afrique et l’Homme. Jacob/Fayard, Paris, 148 p. ISBN 2-213-01272-5
- ↑ Coppens Y., 2006. Histoire de l'homme et changements climatiques. Collège de France/Fayard, 93 p., p. 23. ISBN 2-213-62872-6
- ↑ Patterson N., Richter D. J., Gnerre S., Lander E. S. et Reich D., 2006. Genetic evidence for complex speciation of humans and chimpanzees. Nature, 441 (7097): 1103-1108.
- ↑ Suwa G., Kono R. T., Katoh S., Asfaw B. et Beyene Y., 2007 A new species of great ape from the late Miocene epoch in Ethiopia. Nature, 448 (7156): 921-924. (voir les restes)
Voir aussi
Bibliographie
- Beauvilain Alain, 2003. Toumaï, l’aventure humaine. La Table Ronde, Paris, 239 p. (ISBN 978-2-7103-2592-5)
- Brunet Michel, 2006. D’Abel à Toumaï. Nomade, chercheur d’os. Odile Jacob, Paris, 254 p. (ISBN 978-2-7381-1738-0)
- (en) Gibbons Ann, 2006. The first human.. Doubleday, New York, 306 p. (ISBN 978-0385512268)
- Picq Pascal, 2003. Au commencement était l'homme - De Toumaï à Cro-Magnon. Odile Jacob, Paris, 256 p. (ISBN 978-2-7381-1281-1)
Documentaire
- Toumaï, le nouvel ancêtre, film documentaire et fiction réalisé par Pierre Stine, sous la direction scientifique de Michel Brunet, sorti en 2006.
Articles connexes
- Paléoanthropologie
- hominine
- Autres hominines du Miocène supérieur : Orrorin tugenensis, Ardipithecus kadabba
- Chororapithecus abyssinicus
- chimpanzé
- gorille
Liens externes
- Référence The Paleobiology database : Sahelanthropus tchadensis Brunet et al. 2002 (en)
- « Toumaï l'ancêtre des humains », dossier de presse du CNRS
- Toumaï, l'ancêtre des humains : nouveaux fossiles et reconstruction 3D du crâne, communiqué de presse du CNRS, 7 avril 2005
- Sahelanthropus tchadensis sur Hominidés.com
- Une étude confirme l'âge de Toumaï, site de l'Université de Poitiers
- Toumaï : composition détaillée de la Mission paléoanthropologique franco-tchadienne, de ses missions, des conditions de la découverte de Toumaï ; controverse au sujet du mauvais placement d'une molaire, du nombre minimum d'individus, de la géologie du site ; deux articles contredisant l'hypothèse d'un hominidé bipède
- (en) Toumaï, site au contenu similaire au site précédent mais en anglais
- Toumaï, une femelle protogorille ?, transcription d'un article de Sciences et Avenir d'août 2002
- Photographie de la SEPULTURE de TOUMAÏ
- Photographie du FEMUR de TOUMAÏ
- pages permettant une comparaison visuelle des crânes de Toumaï et d'un gorille femelle :
- Toumaï, la plus ancienne espèce connue d’hominidé
- Toumaï, compte rendu de la conférence donnée par Patrick Vignaud le 15 janvier 2003, au lycée Jean Lurçat de Martigues
- site de la cité des sciences :
- Toumaï le trouble-fête, interviews sonores des grands spécialistes français au sujet de Toumaï
- Toumaï, suite et fin de la controverse ?

La version du 20 juin 2008 de cet article a été reconnue comme « article de qualité », c'est-à-dire qu'elle répond à des critères de qualité concernant le style, la clarté, la pertinence, la citation des sources et l'illustration.  Portail de la paléontologie
Portail de la paléontologie Portail de la Préhistoire
Portail de la Préhistoire Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail de la zoologie
Portail de la zoologie
Catégories : Article de qualité | Hominidé disparu | Recherche paléoanthropologique au Tchad








Wikimedia Foundation. 2010.